Restez toujours informé: suivez-nous sur Google Actualités (icone ☆)
La formation programmée de nombreuses cassures de l'ADN lors de la méiose est un phénomène particulièrement étonnant dont la compréhension moléculaire et évolutive est d'une importance majeure en biologie. L'équipe de Bernard de Massy à l'Institut de génétique humaine à Montpellier, en collaboration avec Henri-Marc Bourbon au Centre de biologie du développement à Toulouse, révèle qu'une protéine essentielle pour la formation de ces cassures est homologue à une sous-unité de topoisomérase. Ceci démontre que les protéines qui catalysent les cassures de l'ADN en méiose ont une activité biochimique en partie similaire à celle des topoisomérases et met en évidence l'évolution d'un ensemble moléculaire qui semble s'être différencié avec l'émergence de la reproduction sexuée. Cette étude est publiée dans la revue Science.
Chez tous les organismes à reproduction sexuée, des cellules reproductrices haploïdes sont générées à partir de cellules diploïdes lors d'une étape de division spécialisée appelée méiose. Lors des premières étapes de la méiose, la recombinaison homologue permet des échanges entre chromosomes parentaux. La recombinaison en méiose a des conséquences biologiques majeures puisque d'une part son absence conduit à la stérilité et que d'autre part elle augmente la diversité à chaque génération et est ainsi un moteur de l'évolution.
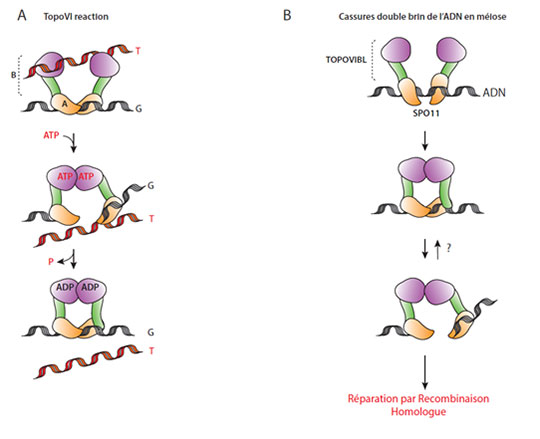
Figure: A. Schéma de la réaction de coupure et religation de la topoisomérase TopoVI: La sous-unité A (TopoVIA) possède l'activité catalytique de coupure de l'ADN (double hélice G) qui requiert aussi la sous-unité B (TopoVIB). TopoVIB possède un domaine de fixation de l'ATP (rose) et un domaine d'interaction avec la sous-unité A (vert). Les interactions entre les sous-unités B et la fixation et l'hydrolyse de l'ATP permettent la coupure et le passage de la double hélice T à travers la molécule G coupée. B. Formation de coupure de l'ADN en méiose: SPO11, orthologue de TopoVIA, possède l'activité catalytique de coupure de l'ADN. Cette coupure requiert la sous unité TOPOVIBL qui interagit avec SPO11. Il est possible, mais non démontré, que TOPOVIBL fixe l'ATP et/ou dimèrise. L'ADN coupé est réparé par la recombinaison homologue mais pourrait dans certains contextes se recoller.
© Thomas Robert
Réaliser des échanges entre chromosomes nécessite de couper et recoller les brins de la double hélice d'ADN. Ces réactions sont effectuées en plusieurs étapes qui ont été en partie décrites au niveau moléculaire chez des organismes modèles et de nombreux travaux de recherche actuels visent à identifier les protéines qui réalisent ces échanges.
Une observation remarquable, datant de plus de 15 ans, a permis de démontrer qu'une protéine directement impliquée dans les coupures de l'ADN en méiose, Spo11, est phylogénétiquement reliée à des enzymes de la famille des topoisomérases appelées TopoVIA. Ces enzymes, d'abord identifiées chez les archaebactéries qui n'ont pas de cycle de reproduction sexuée, ont pour fonction de couper et recoller l'ADN et ainsi d'en réguler la topologie . Ces données sont fascinantes puisqu'elles suggèrent une évolution moléculaire qui aurait conduit à la modification d'une activité enzymatique impliquée dans le contrôle de la topologie de l'ADN pour promouvoir des échanges entre chromosomes. Dans ce contexte, une question centrale a été de comprendre cette évolution et en particulier de déterminer si la protéine Spo11 agit en interaction avec une autre sous-unité, à l'instar de ce qui est décrit dans la famille des TopoVI où TopoVIA interagit avec TopoVIB. Malgré de multiples tentatives, ces recherches n'avaient pas abouti jusqu'à récemment. Grâce à un crible génétique, l'équipe de Mathilde Grelon à l'Institut Jean-Pierre Bourgin (INRA-AgroParisTech) a identifié un gène d'Arabidopsis thaliana dont la mutation conduit à l'absence de coupure de l'ADN en méiose, au même titre qu'un mutant dans le gène Spo11 .
A partir de la séquence de ce gène, l'équipe de Bernard de Massy, en collaboration avec Henri-Marc Bourbon, a recherché des orthologues dans d'autres espèces et abouti à deux conclusions majeures: cette protéine est conservée chez de nombreuses espèces au sein des opisthokonta (animaux et champignons), et elle est homologue à la sous-unité de topoisomérases d'archaebactéries TopoVIB. Les chercheurs, ayant ainsi potentiellement découvert la deuxième sous-unité de Spo11, ont proposé de l'appeler TOPOVIB-Like (TOPOVIBL). La particularité de cette similarité entre TOPOVIB-Like et TopoVIB est qu'elle repose sur une très faible conservation au niveau de la séquence primaire en acides aminés (11% d'identité), ce qui explique qu'elle était restée non détectée jusqu'à ce jour. En revanche, les chercheurs ont pu mettre en évidence une forte conservation au niveau de la structure secondaire et tertiaire.
Afin de valider la fonction de cette nouvelle famille de protéines, TOPOVIB-Like, les chercheurs ont développé une étude moléculaire et fonctionnelle de la protéine murine qui avait pour but de vérifier son interaction avec SPO11 et son rôle dans la formation des cassures de l'ADN en méiose. Ils ont démontré que TOPOVIBL interagissait et formait un complexe avec SPO11 puis ils ont introduit une mutation pour invalider le gène murin (Top6bl) par une délétion générée par CrispR/Cas9. Chez les souris homozygotes Top6bl-/Top6bl-, les analyses de spermatocytes et d'ovocytes montrent l'absence de cassure de l'ADN en méiose de manière similaire à ce qui est observé chez un mutant dans le gène Spo11. La méiose et donc la formation de cellules reproductrices mâles et femelles sont altérées, les mâles ne produisent aucun spermatozoïde, et les souris sont stériles.
Au niveau évolutif, il apparait que c'est l'ensemble d'un complexe protéique de type topoisomérase qui a été modifié, ce qui a conduit à l'acquisition d'une activité spécifique lors de la méiose. Au niveau moléculaire, la présence de cette deuxième sous-unité suscite immédiatement une question: la formation des cassures de l'ADN en méiose est-elle réversible comme elle l'est dans le cas des cassures induites par les topoisomérases ? Cette question est conceptuellement liée à celle de la régulation de l'induction des cassures de l'ADN en méiose: comment contrôler une machinerie enzymatique qui induit la formation de plusieurs centaines de cassures de l'ADN dans le génome et qui doivent être réparées sans erreur? Des analyses plus approfondies sur les protéines TOPOVIB-Like, leurs structures et leur diversité, devraient permettre d'identifier ces mécanismes de régulation aujourd'hui inconnus.
Populaires