Protéine du rétinoblastome - Définition
La liste des auteurs de cet article est disponible ici.
Régulation de la fonction biologique
Sous l’influence de facteurs mitogènes (par exemple PDGF) les cellules parviennent à lever le frein biologique posé par la présence de pRB hypophosphorylé et ainsi continuer la progression du cycle cellulaire vers la mitose.
Suite à la liaison du facteur mitogène et de son ligand, une cascade de signalisation est enclenchée. Cette cascade de signalisation utilise des kinases pour phosphoryler le substrat suivant de la cascade jusqu’à l’activation du facteur de transcription. Une fois le facteur de transcription activé, cela enclenche la production de protéines nommées cyclines et CDKs, nécessaires pour démarrer le cycle cellulaire.
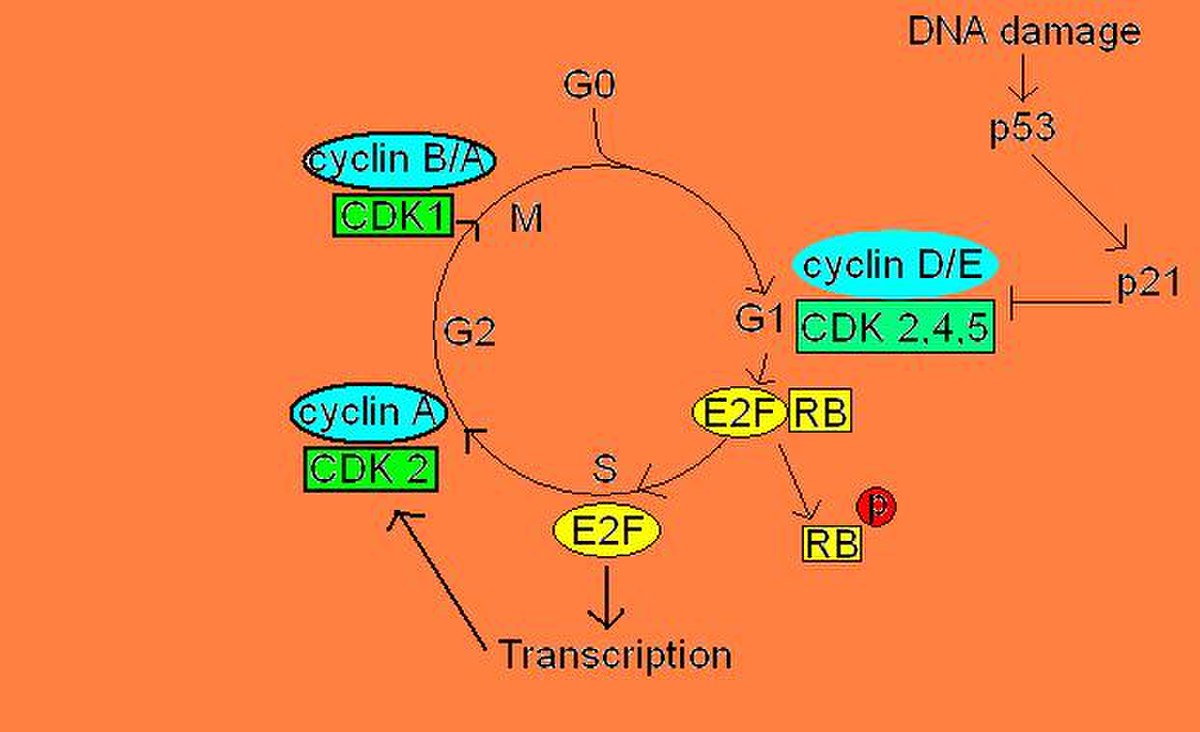
Les cyclines et CDKs nécessaire pour la phase G1, ce qui veux dire qu’ils sont essentiels pour arriver à la phase S, sont les cyclines D et E et les CDKs 2/4/6. Cependant, la cycline D et CDKs 4 et 6 sont produite et activées au tout début de la phase G1, mais ne sont pas capable d’induire la libération du complexe E2F/DP. En effet, on doit attendre à la fin de la phase G1 pour voir la production de cycline E activer CDK 2 qui, à son tour phosphorylera le domaine carboxyl-terminal de pRB. Cette phosphorylation en chaîne par les trois CDKs entraîne un changement de conformation dans la structure de pRB, ce qui l’empêche alors de lier le complexe E2F/DP. Ce complexe étant maintenant libre, le facteur de transcription E2F peut alors aller initier la transcription d’ARNm nécessaires à la phase S du cycle cellulaire.
Cependant, il peut aussi y avoir une régulation négative de pRB. En effet, s’il y a des dommages à l’ADN lorsque la cellule veut entrer en mitose, la protéine p53 se trouve phosphorylée et induit la production de protéine p21. p21 se lie aux complexes cycline/CDK de la phase G1, inhibant ainsi leur activité kinase pour pRB. Il y a donc inhibition de la libération du complexe E2F/DP.
Pathologies fréquemment associées
- Rétinoblastome (sporadique unilatéral ou héréditaire bilatéral) ;
- Tumeur de Wilms ;
- Ostéosarcome ;
- Cancer de la vessie.

















































