GroEL - Définition
La liste des auteurs de cet article est disponible ici.
Introduction
La protéine GroEL appartient à la famille des chaperonines des molécules chaperonnes, et se trouve chez un grand nombre de bactéries. Elle est nécessaire pour le repliement efficace de nombreuses protéines. Afin de fonctionner efficacement, GroEL requiert le complexe protéique couvercle associé GroES. Chez les eukaryotes, les protéines Hsp60 et Hsp10 sont structurellement et fonctionnellement presqu'identiques à GroEL et GroES, respectivement.
Mécanisme d'action
Dans une cellule, le processus de repliement de protéine par médiation GroEL/ES implique de nombreuses séquences de liaisons, encapsulations et libération de protéine substrat. Les protéines substrats non repliées se lient à une aire hydrophobe sur le bord intérieur de la cavité ouverte du GroEL, formant ainsi un complexe binaire avec la chaperonine. La liaison de protéine substrat de cette façon, en plus de la liaison d'ATP, conduit à une modification conformationnelle qui permet l'association du complexe binaire avec une structure couvercle distincte, GroES. La liaison de GroES avec la cavité ouverte de la chaperonine provoque la rotation des sous-unités individuelles de la chaperonine de manière à ce que le substrat hydrophobe liant soit soustrait à l'intérieur de la cavité, ce qui induit l'éjection de la protéine substrat du bord vers la chambre maintenant hydrophile. L'environnement hydrophile de la chambre favorise l'enfouissement des résidus hydrophobes du substrat, ce qui induit le repliement du substrat. L'hydrolyse de l'ATP et la liaison d'une nouvelle protéine substrat dans la cavité opposée envoie un signal allostérique qui provoque la libération de GroES et d'une protéine encapsulée dans le cytosol. Une protéine donnée subira plusieurs cycles de repliement, retournant à chaque fois à son état non-replié original, jusqu'à ce que la conformation native ou une structure intermédiaire destinée à atteindre l'état natif soit produite. De manière alternative, le substrat peut être dénaturé lors d'une réaction compétitive, comme un mauvais repliement ou une agrégation avec d'autres protéines mal repliées.
Thermodynamique de la réaction
La nature contractée de l'intérieur du complexe moléculaire favorise fortement les conformations moléculaires compactes de la protéine substrat. En solution, les interactions apolaires et à longues portées peuvent seulement se produire dans ce cas avec un coût important en entropie. Dans les espaces fermés du complexe GroEL, la perte relative d'entropie est beaucoup plus faible. La méthode de capture tends aussi à concentrer les sites liants apolaires séparément des sites polaires. Lorsque les surfaces apolaires de GroEL sont ôtées, la chance pour qu'un quelconque groupe apolaire donné puisse rencontrer un site apolaire intramoléculaire sont bien plus importantes qu'en solution. Les sites hydrophobes se situant à l'extérieur sont rassemblés au sommet du domaine cis et liés entre eux. La géométrie de GroEL impose que les structures polaires soient motrices, et elles enveloppent le cœur apolaire lorsqu'il émerge depuis le côté trans.
Interactions
Il a été montré que GroEL interagit avec GroES,, l'ALDH2, la caspase 3, et la reductase dihydrofolate.
Structure
GroEL est, structurellement, un tétradécamère à anneaux doubles, les anneaux cis et trans étant constitués de sept sous-unités chacun. L'intérieur de GroEL est hydrophobe, et constitue l'endroit où le repliement de protéine est censé se faire.

GroEL (côté) | 
GroEL (dessus) | 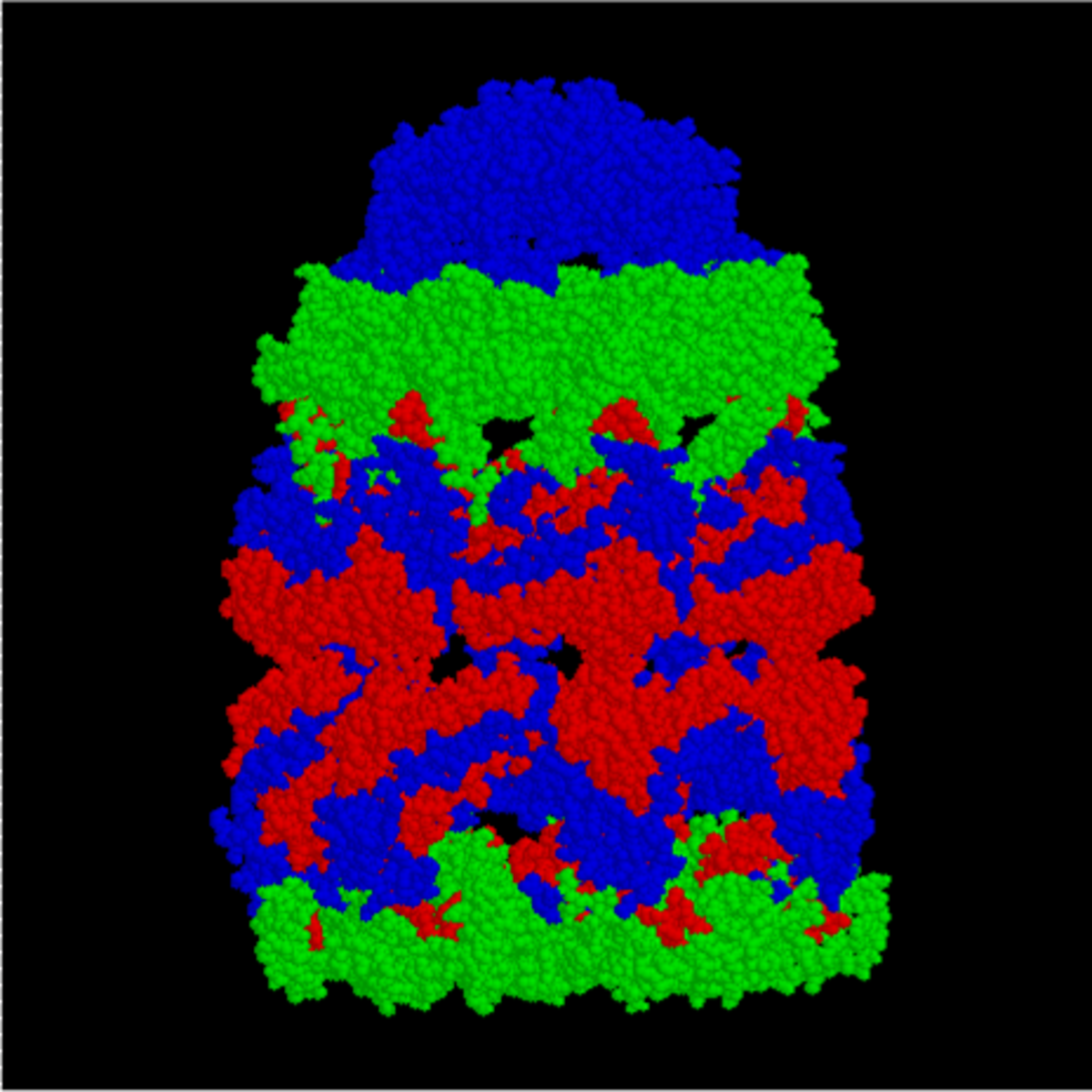
Complexe GroES/GroEL (côté) | 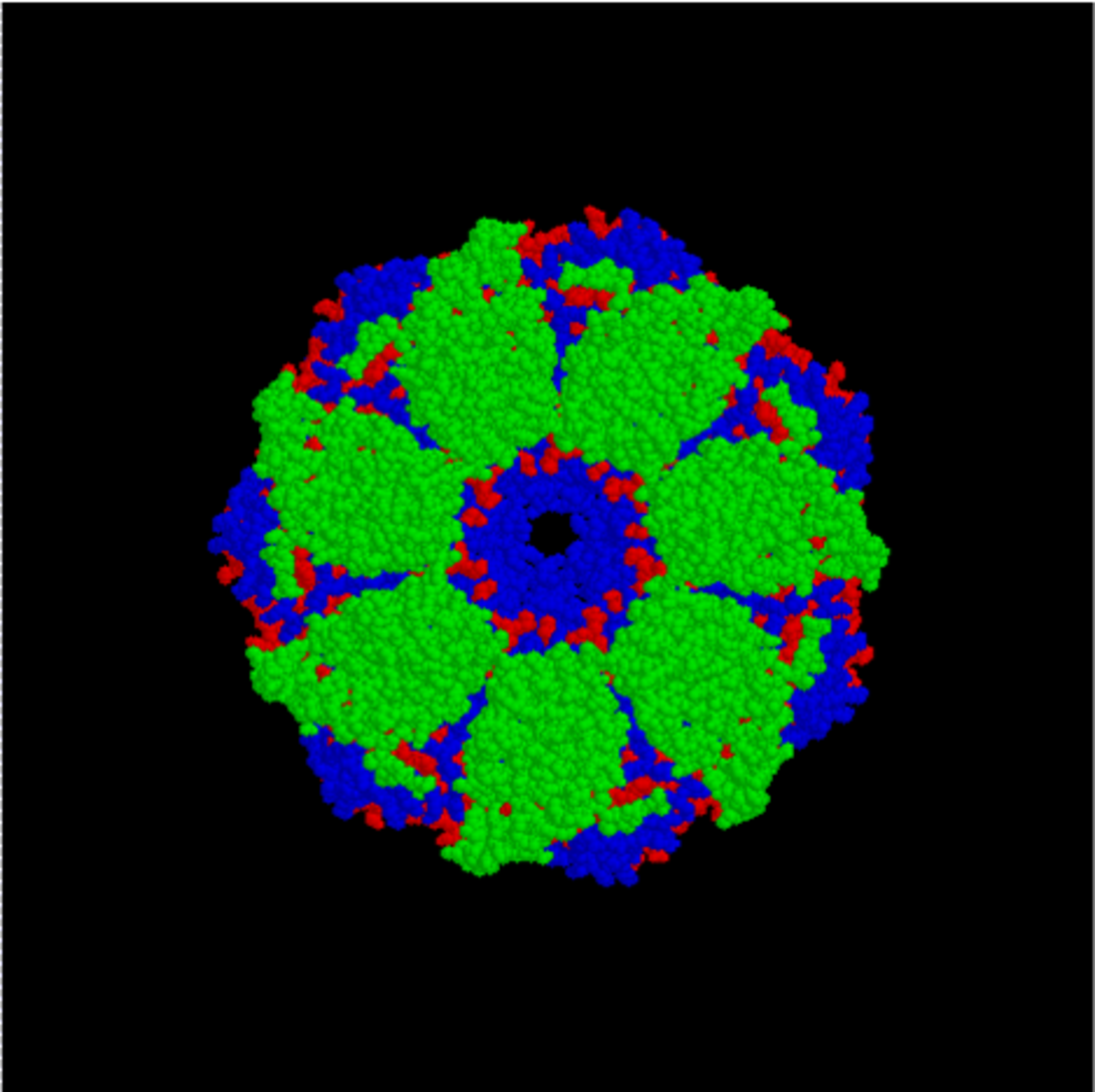
Complexe GroES/GroEL (dessus) |
La clé de l'activité de GroEL réside dans la structure du monomère. Le monomère Hsp60 possède trois parties distinctes séparées par deux régions charnières. La région apicale contient un grand nombre de sites liants pour les substrats pour protéines non repliées. De nombreuses protéines globulaires ne se lieront pas à cette région apicale car leurs parties hydrophobes sont internes, loin du milieu aqueux, dans une conformation favorisée par la thermodynamique. En conséquence, ces « sites substrats » ne se lieront qu'avec des protéines qui ne sont pas repliées de manière optimale. La région apicale possède aussi des sites liants pour les monomères Hsp10 de GroES.
Le domaine équatorial possède un emplacement près du point charnière pour lier l'ATP, ainsi que deux points de rattachement pour l'autre moitié de la molécule GroEL. Le reste de la région équatoriale est modérément hydrophile.
L'ajout de l'ATP et de GroES induit un effet drastique sur la conformation de la région cis. Il est causé par le pliement et la rotation des monomères Hsp60 aux points charnières. La région intermédiaire se replie vers l'intérieur, d'environ 25° par rapport à la charnière la plus basse. Cet effet, multiplié par le pliement coopératif des monomères, accroît le diamètre équatorial de la cage GroEL. Mais le domaine apical pivote de 60° vers l'extérieur par rapport à la charnière suppérieure, et aussi de 90 ° autour de l'axe charnière. Ce mouvement ouvre très largement la cage au dessus de la région cis, mais supprime l'ensemble des sites liant de l'intérieur de la cage.

















































