Sélection intersexuelle - Définition
La liste des auteurs de cet article est disponible ici.
Bénéfices et coûts du choix de la paternité
Chez de nombreuses espèces sexuées, les femelles exercent un choix sur les mâles qui porteront la paternité de leur descendants, exerçant ainsi une pression de sélection intersexuelle sur ces mâles. Ces choix, qui détermineront les caractères sélectionnés chez les mâles, peuvent eux-mêmes avoir été sélectionnés parce qu'ils augmentent le fitness des femelles, ou peuvent être apparus par hasard sans apporter d'avantage sélectif aux femelles. Par ailleurs, le choix du ou des mâles qui seront les pères de leurs descendants a souvent un coût pour les femelles, généralement inférieur ou égal à l'avantage sélectif conféré par ce choix.
Avantage sélectif du choix
Le fait pour une femelle de choisir les mâles qui lui donneront des descendants, selon certains critères particuliers, peut avoir été sélectionné au cours de l'évolution, car ce choix confère un avantage sélectif aux femelles, celles choisissant leurs mâles ayant plus de descendants viables et fertiles. Les avantages sélectifs que peuvent conférer un choix sont divers et nombreux. Ainsi, le bénéfice obtenu par la femelle peut être direct, et se présenter sous forme de nourriture apportée à la femelle, ou encore de soins parentaux conférés par le mâle. Les bénéfices peuvent aussi être indirects, sous la forme de gènes capables d'augmenter le fitness de la femelle parce qu'ils produiront des descendants plus viables (parce qu'ils seront plus attrayants pour les autres femelles ou plus aptes à survivre par exemple).
Bénéfices directs
Le fait de choisir les mâles peut conférer un bénéfice direct aux femelles si les critères de ce choix reflètent la capacité du mâle à fournir divers avantages matériels. Ceux-ci peuvent-être des dons nutritifs, comme des proies chassées, des nutriments synthétisés par le mâle, voir le mâle lui-même qui s'offre à être mangé si le bénéfice de ce sacrifice est suffisamment grand pour le mâle. Les femelles peuvent aussi avoir avantage à choisir le mâle selon des critères permettant d'évaluer sa capacité à fournir des soins aux enfants, sous forme de nutrition ou de protection par exemple. Les mâles sont parfois choisis parce qu'ils sont plus aptes à défendre les femelles, ou parce qu'ils gardent des territoires plus avantageux (plus riches en ressources par exemple). Le choix du mâle sur des critères reflétant la fécondité du mâle peut aussi garantir à la femelle que ses ovules seront bien fécondés. Enfin, les signaux mâles peuvent garantir la bonne santé du mâle et éviter à la femelle la contamination par des parasites lors de la fécondation.
Avantages nutritifs
Soins parentaux des descendants
Défense d'un territoire
Fécondité du mâle
Risques de contamination et de prédation
Bénéfices indirects
Le bénéfice conféré par le choix du mâle peut aussi être indirect, s'il n'affecte non pas directement le fitness de la femelle, mais celui de ses descendants. Les avantages sont alors transmis sous forme génique par le mâle à ses descendants, et le fitness de la femelle est augmenté indirectement, par le biais de ses descendants. Les avantages génétiques conférés aux descendants peuvent être de plusieurs types. Si la préférence maternelle est déterminée génétiquement, la préférence pour un trait et le trait préféré vont être transmis ensemble à la génération suivante, donnant lieu à un phénomène d'auto-amplification appelé runaway de Fisher. Choisir un mâle considéré comme plus sexy garantit donc aux descendants de la femelle d'avoir des descendants mâles plus à même d'être choisis par les femelles. La femelle peut aussi choisir le mâle selon des critères qui reflètent la qualité génétique de celui-ci, et donc la capacité de survie des descendants, en évaluant par exemple sa capacité à produire des organes coûteux. Enfin, une femelle peut choisir le mâle susceptible de fournir les descendants les plus viables compte tenu de son propre potentiel génétique. L'hétérozygotie étant généralement considéré comme un avantage, la femelle choisit alors un mâle dont le patrimoine génétique est le plus différent du sien, par le biais du complexe majeur d'histocompatibilité par exemple. De même, les femelles peuvent exercer un choix post-copulatoire permettant de sélectionner les gamètes qui sont compatibles avec ses ovules, ce choix pouvant donc être différent d'une femelle à l'autre. Une telle sélection de la compatibilité génétique permettrait aux femelles d'éviter la fécondation par du sperme incompatible, et donc l'avortement de l'embryon, à cause de conflits cytoplasmiques par exemple.
Runaway de Fisher
Indicateurs de la qualité génétique
Complémentarité génétique
Compatibilité génétique
Caractère non-adaptatif
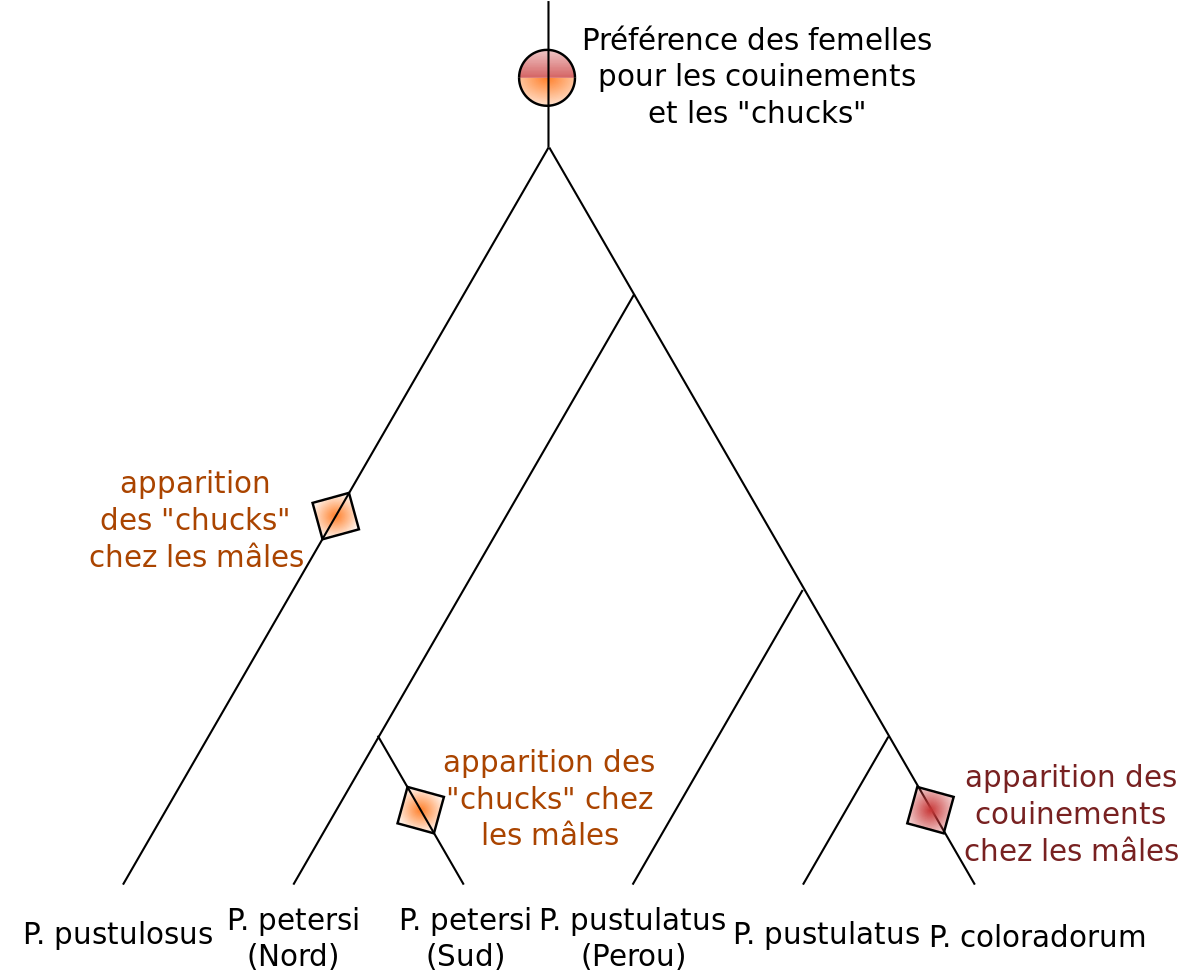
Il est aussi possible que les femelles exercent un choix sur la paternité de leurs descendants sans que ce choix n'ait été sélectionné, et le choix n'apporte aucun bénéfice ou coût supplémentaire notable pour la femelle. La préférence de la femelle peut-être alors due à un biais dans son système sensoriel, sélectionné pour des raisons autres que la reproduction, comme la recherche de nourriture. Les mâles exploitant ce biais sensoriel sont alors préférés et sélectionnés. Cette hypothèse, au point de départ non adaptative, peut-être renforcée par un phénomène de coévolution antagoniste entre les mâles et les femelles. En effet, si le choix engendré par ce biais sensoriel diminue le fitness de la femelle, ce choix va être contre-sélectionné, et la sensibilité de la femelle pour ce caractère peut baisser. Les mâles dont la valeur de ce caractère dépasse ce seuil de sensibilité vont alors être sélectionnés, favorisant l'évolution d'une baisse de sensibilité encore plus importante chez les femelles.
Exploitation d'un biais sensoriel
Coévolution antagoniste entre les sexes
Coûts du choix de la paternité
Le choix des femelles a aussi un coût, qui peut ne pas être négligeable. Ce coût peut prendre différentes formes. Tout d'abord, le choix lui-même est coûteux. Premièrement en termes de temps, la femelle ne pouvant se consacrer à d'autres activités telles que la recherche de nourriture ou le soin des petits pendant ce temps. Il est aussi coûteux en énergie, laquelle est nécessaire à l'échantillonnage des mâles ou des gamètes et à la prise de décision elle-même. De plus, le choix peut engendrer des coûts indirects, par exemple si la femelle est plus vulnérable aux prédateurs pendant la période de choix.

















































