Microsporidia - Définition
La liste des auteurs de cet article est disponible ici.
Introduction
| |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Classification phylogénétique | |||||||||
| Domaine : Eukaryota | |||||||||
| Taxons de rang inférieur | |||||||||
| |||||||||
| Classification classique | |||||||||
| Fungi | |||||||||
| | |||||||||
Les microsporidies sont des eucaryotes, parasites intracellulaires obligatoires, classés parmi les Mycètes. Il semblerait qu'ils soient génétiquement très proches des Eumycètes dont ils dérivent peut-être par adaptation au parasitisme. La comparaison des régions de synténie avec d'autres champignons montre que les microsporidies seraient proches des Mucoromycotina. On en connait maintenant 150 genres et 1200 espèces. Cependant, une nouvelle estimation du nombre d’espèces montre qu’il se pourrait que le nombre d’espèce de microsporidies atteigne le nombre d’espèces animales.
Les microsporidies présentent une adaptation caractérisée par la réduction. En effet les microsporidies sont réduites à beaucoup de niveaux par rapport aux autres eucaryotes ; au niveau du génome en premier lieu mais aussi au niveau de leur métabolisme.
Le génome de Encephalitozoon cuniculi a été entièrement séquencé. Ce parasite possède le génome eucaryote le plus petit connu à ce jour.
Histoire
La première description d’une microsporidie fut faite au milieu du 19ème siècle, quand la pébrine décimait les vers à soie. Cette maladie causée par Nosema bombycis fut identifiée par Karl Wilhelm von Nägeli en 1857.. Ce champignon semble avoir été signalé pour la première fois en 1849, avant l'identification de la pébrine, par Guérin-Méneville, qui croyait erronément que les vers à soie sur lesquels il l'observait étaient atteints de muscardine. Après l'identification de la pébrine, Cornalia, Franz Leydig, Balbiani et Pasteur étudièrent son agent microbien (« corpuscules ») de façon plus approfondie. C’est ensuite Louis Pasteur qui eut l’idée de sélectionner les individus non infectés pour recréer de nouveaux élevages sains.
Structure
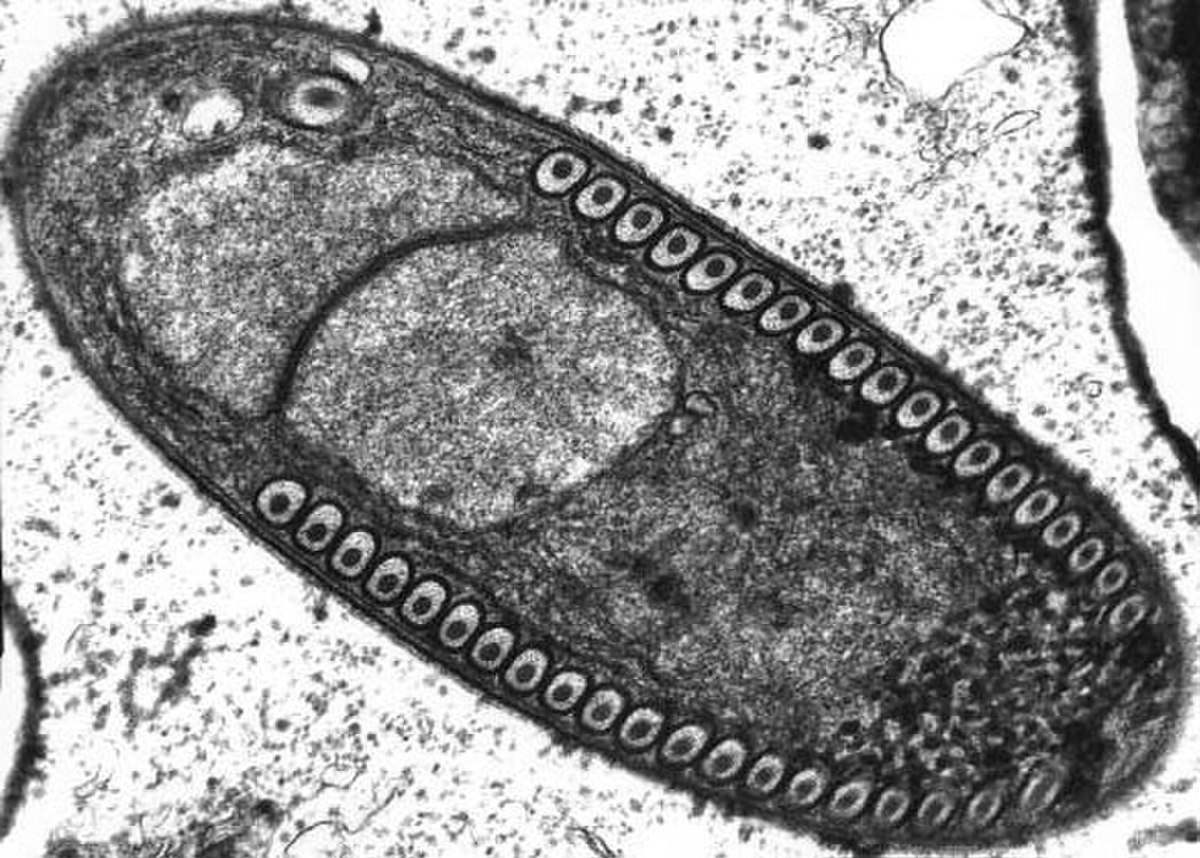
Les microsporidies présentent une forme de résistance et de dissémination : la spore. La spore est le seul stade reconnaissable des microsporidies. C’est à ce stade que les espèces peuvent être différenciées et c’est le seul à être viable hors de la cellule hôte. La taille et la forme de la spore sont très conservés chez une même espèce.
La spore est entourée par une membrane plasmique classique ainsi que par deux parois extracellulaires rigides : exospore et endospore. L’exospore et faite d’une matrice glycoprotéique et fibreuse dense. L’endospore est composée de chitine alpha et d’autres protéines. Son épaisseur est assez uniforme sauf à l’apex où cette paroi est plus fine. À l’intérieur de la membrane se trouve le sporoplasme (cytoplasme de la spore) qui constitue le matériel infectieux. Il contient un seul noyau, il est riche en ribosomes et est occupé majoritairement par les trois structures nécessaires à l’infection:
- le polaroplaste
- le filament polaire ou tube polaire
- la vacuole postérieure
Le polaroplaste est une grande organisation de membrane occupant la partie antérieure de la spore. La portion antérieure du polaroplaste est hautement organisée sous forme de membranes empilées appelées polaroplaste lamellaire alors que la portion postérieure est moins organisée et est appelée polaroplaste vésiculaire. L’organite jouant le rôle le plus évident dans l’infection est le filament polaire (ou tube polaire). Dans le sporoplasme, il est composé de couche de glycoprotéines ; 0.1 à 0.2 µm de diamètre et 50 à 500 µm de long. Il est attaché à l’apex via une structure en parapluie appelée disque d’ancrage. Sur 1/3 de la spore ce filament est raide et hélicoïdal (le nombre de spires et leur angle sont conservés et permettent d’identifier certaines espèces). Ce filament se termine au niveau de la vacuole postérieure. Il semblerait qu’il y ait un contact physique entre ces 2 structures.

















































