Système du complément - Définition
La liste des auteurs de cet article est disponible ici.
Voie effectrice commune
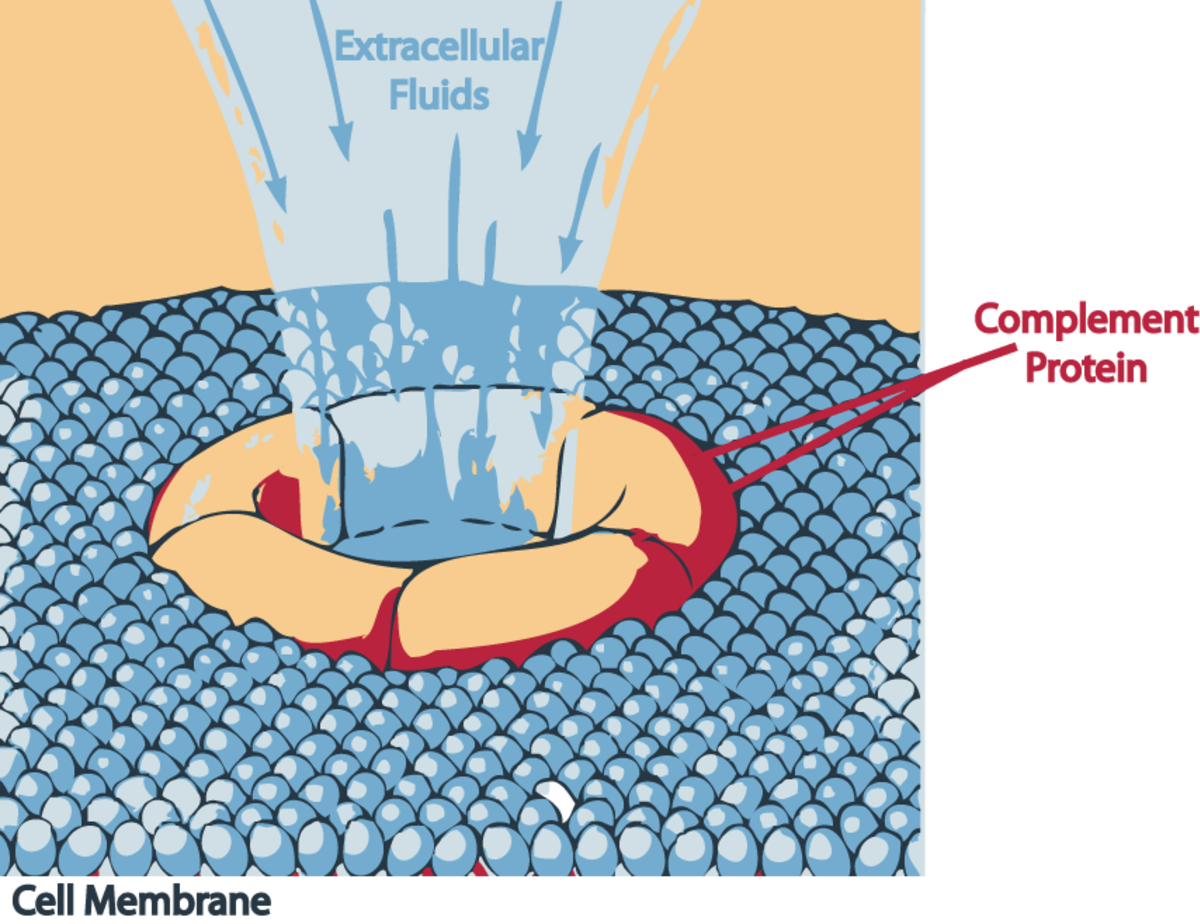
Dans les deux cas, nous verrons la voie alterne ensuite, les composants précoces activent localement C3, qui est le facteur pivot du complément, et dont le clivage conduit non seulement à l'assemblage du complexe qui attaque la membrane, mais aussi au recrutement des différents globules blancs.
Le C3b, qui est le plus grand fragment de la lyse de C3 par la C3-convertase, se lie de façon covalente à la surface de la cellule. Le plus petit fragment, C3a, agit quant à lui comme signal diffusible qui provoque une réponse inflammatoire, en stimulant la migration des globules blancs vers le site de l'infection. Le fragment C3b fixé à la membrane, produit à la fois par la voie classique et la voie alterne, et même la voie des lectines, amorce la cascade des réactions qui conduit à la formation du complexe d'attaque membranaire, à partir des composants tardifs du complément. Il est donc fixé sur la membrane de façon covalente, et il clive le facteur C5 en C5a et C5b. C5b reste faiblement lié à C3b et s'assemble rapidement à C6 et C7 pour former le complexe C567 qui va s'ancrer à la membrane via C7. Ce complexe lie ensuite C8, pour former le complexe C5678. La liaison du facteur C9, qui expose une région hydrophobe après changement de conformation, entraîne son insertion dans la membrane plasmique de la cellule. Il s'ensuit alors une réaction en chaîne où les C9 de nouvelle conformation vont lier des C9 de l'ancienne, entraînant le changement conformationnel qui leur permet de s'insérer dans la double couche lipidique. C'est ainsi qu'il se forme un canal au travers de la membrane cellulaire. Dès lors, la perméabilité de la cellule est perturbée, les petites molécules pénètrent et sortent de la cellule au voisinage de ces pores, et à travers ceux-ci. Les macromolécules ne peuvent cependant pas passer. De ce fait, le mécanisme cellulaire contrôlant l'équilibre des échanges est bouleversé. L'eau entre par osmose dans la cellule, faisant augmenter son volume jusqu'à la lyse. On observe ce même phénomène de lyse avec des globules rouges en solution hypotonique. Ce système est très efficace puisqu'il a été observé que la présence d'un seul de ces canaux permet la lyse d'un globule rouge.
Rôle dans les maladies
Les maladies liées à un déficit en complément sont rares :
- les déficits en protéines de la voie classique : C1q, C2, C4, se révèlent par un lupus
- les déficits en protéines de la voie des mannanes : MBP, donnent des infection graves et récidivantes
- les déficits en protéines de la voie alterne : B, D, Properdine, donnent des infections bactériennes graves
- les déficits en protéines du complexe d'attaque membranaire : C5, C6, C7, C8, s'accompagnent d'infections à Neisseria
- les déficits en inhibiteurs : H, I donnent des infections avec urticaire, anémie hémolytique, glomérulopathie ou les déficits en CD55 et CD59 donnent l'Hémogloginurie Paroxystique Nocturne (maladie de Marchiafava et Michelli) ou encore les déficits en C1inh (œdème angio-neurotique).
- des déficits en CR1 et CR3 ont aussi été décrits.
Les maladies accompagnées de dysfonctionnements du complément sont nombreuses.
Bien que quelques agents pathogènes soient capables d'inactiver le complément, la plus grande majorité ne le peut pas, et au contraire, active la voie alterne de la C3 convertase. Ces agents pathogènes favorisent la production des molécules de la voie alterne d'activation, en particulier C3a, C3b, C5a et le complexe d'attaque membranaire C56789. Celui-ci étant formé sur le site de la C3-convertase, son activité de dégradation s'exerce sur les pathogènes et non sur les cellules de l'hôte. Les fragments solubles du complément C3a et C5a qui sont formés possèdent des propriétés pro-inflammatoires. Par exemple, la liaison du C5a aux récepteurs des cellules endothéliales augmente la perméabilité vasculaire et l'infiltration des protéines plasmatiques dans les tissus inflammés.

















































