Système du complément - Définition
La liste des auteurs de cet article est disponible ici.
Généralités
Le système du complément est un ensemble de protéines circulantes ou membranaires du sang, principalement sécrétées par le foie. Leur rôle initialement reconnu était de compléter l'action des immunoglobulines sériques, d'où leur nom. En l'absence des protéines thermolabiles (qui perdent leur qualités à une température déterminée), les immunoglobulines thermostables spécifiques sont incapables d'entraîner la lyse de leur cible. Les protéines du complément représentent environ 5% des globulines plasmatiques. Les différentes protéines du complément sont des pro-enzymes inactives et qui sont activées en cascade par clivage. Le clivage libère une fraction ayant une activité enzymatique de protéase, et un petit fragment qui possède souvent un rôle sur les cellules inflammatoires.
Le système du complément possède plusieurs fonctions importantes: la cytolyse d'une cellule ou d'un agent pathogène, l'activation du système immunitaire par les petits fragments de clivage pro-inflammatoires, l'opsonisation de certains agents permettant leur phagocytose, et le métabolisme des complexes immuns circulants grâce aux récepteurs des fragments du complément.
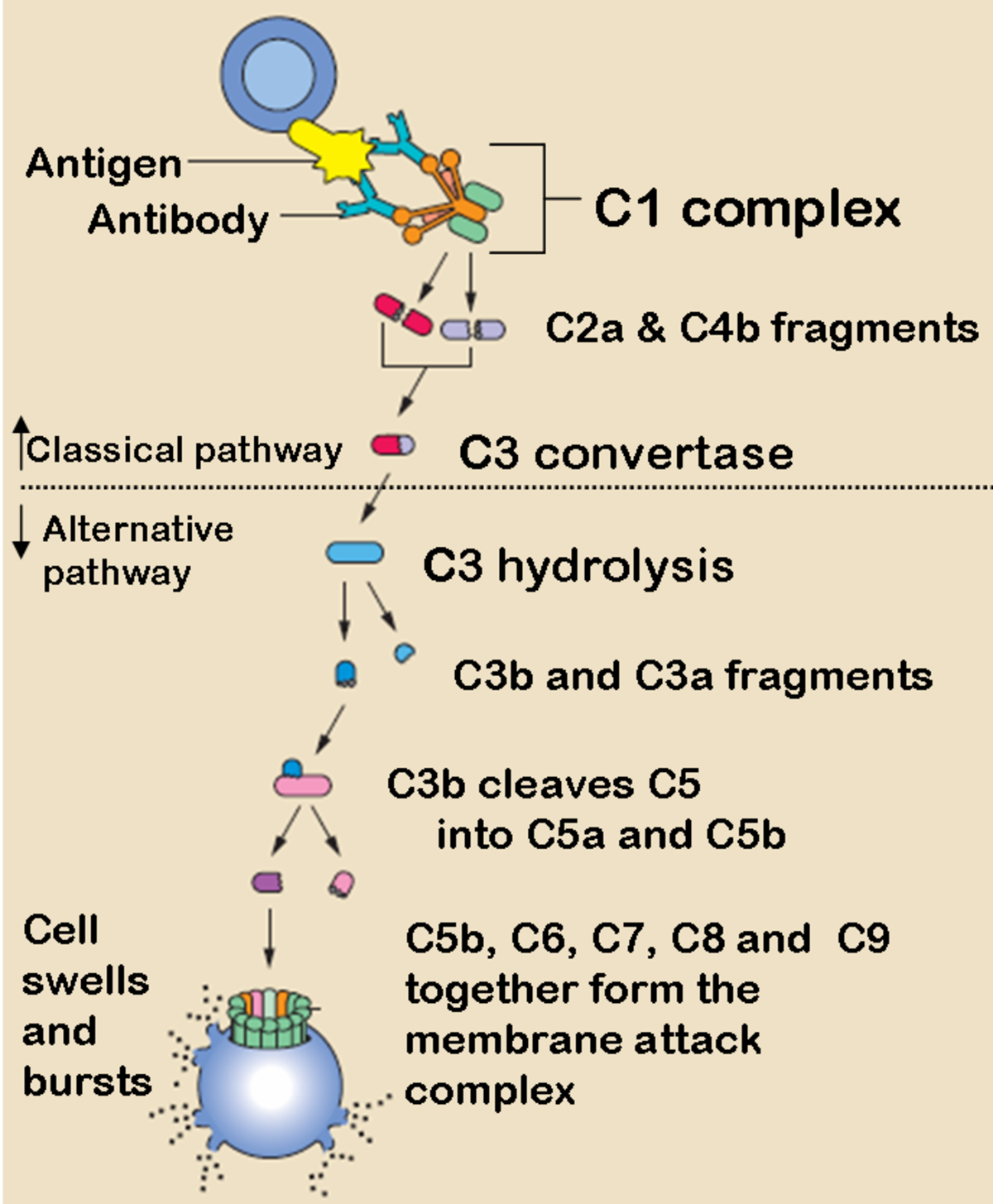
Les différentes voies activant le complément aboutissent à la formation d'une C3 Convertase, point de départ de la voie effectrice commune qui détruit la cible en formant un canal transmembranaire, permettant l'entrée de molécules d'eau dans la cellule. Les principales protéines du complément sont notées de C1 à C9, elles migrent en électrophorèse dans la fraction des Beta Globulines et ont un poids moléculaire de 100 à 200 kDa.
Il existe deux voies principales: La voie alterne et la voie classique, C3 étant le pivot de chacune des voies.
Les fragments C3a et C5a sont des molécules appelées anaphylatoxines: elles ont pour rôle de libérer l'histamine, de plus C5a à une activité chimiotaxique importante. C5a peut aussi activer la voie des lipooxygénases et augmenter l’adhésion leucocytaire. Tandis que C3b et C3bi fonctionnent comme des opsonines.
Les protéines de C5 à C9 en s'assemblant forme le "complexe d'attaque membranaire" (appelée en anglais, le membrane attack complex ou MAC), qui est en l'occurrence l'élément permettant la lyse des cellules pathogènes.
La Voie alterne
La voie alterne est la première défense apparue au cours de l'évolution et est la première en jeu lors d'une infection par un germe inconnu de l'organisme infecté avant une réponse immune spécifique.
Activation
Elle est activée quant à elle par les surfaces cellulaires des bactéries gram+ ou gram-, quelques cellules infectées par un virus, quelques levures, et parasites. Également par les polysaccharides, comme le zymosan ou l'inuline, par les LPS (lipopolysaccharides) bactériens, et diverses substances, comme les fibres d'amiante, le gluten, l'hémoglobine, certains produits de contraste fortement iodés et quelques cellules tumorales. À noter qu'une membrane cellulaire est d'autant plus activatrice de la voie alterne qu'elle est pauvre en acide sialique. La voie alterne d'activation résulte de la fixation du C3b sur un site accepteur. En permanence de faibles quantités de C3 sont clivées spontanément en C3a et C3b. Ce dernier possède, pendant un très court instant, un site hautement réactif capable de se fixer sur des groupements chimiques présents sur virtuellement toutes les surfaces biologiques, principalement bactériennes. En l'absence de ce site accepteur, le C3b réagit avec l'eau et donne le C3b soluble. Le C3b fixé à une surface peut alors lier le facteur B qui est clivé, en présence d'ions Mg2+, par le facteur D en Bb et Ba, formant la C3-convertase alterne, ou C3bBb. Cette C3-convertase clive des molécules de C3 pour former un complexe C3bBbC3b, ou C5-convertase alterne.cette dernière va à son tour cliver le c5 rejoignant ainsi la voie classique.
Régulation
Elle s'effectue grâce à différentes protéines :
- le facteur H qui agit sur la dissociation de la C3/C5 convertase alterne et comme cofacteur du facteur I dans l'inactivation du C3b en C3bi. Le facteur H intervient pour contrôler l'activation de la voie alterne en phase fluide.
- le facteur I qui inactive le C3b en C3bi puis en C3c et C3dg, en présence de facteur H.
- la properdine ou facteur P qui stabilise le complexe C3bBbC3b.
Au niveau des membranes, la régulation de l'activation de la voie alterne se fait grâce à différentes protéines membranaires: le récepteur du C, CR1, le facteur d'accélération de dissociation (DAF), la protéine cofacteur de membrane (MCP) ou CD46. Ces protéines agissent comme cofacteur du facteur I pour dégrader C3b et C4b ou comme accélérateur de la dissociation des C3/C5 convertase alterne ou classique (CR1 et DAF).

















































