Vol de l'insecte - Définition
La liste des auteurs de cet article est disponible ici.
Mécanisme de vol indirect

Le mécanisme de vol indirect n'est manifestement apparu qu'une seule fois au cours de l'évolution, chez l'ancêtre de tous les autres insectes volants. C'est le caractère commun, ou synapomorphique, qui permet de définir l'infraclasse des néoptères : les muscles responsables du vol ne sont pas directement attachés aux ailes.
Ce caractère est associé au mécanisme de repliement des ailes sur le dessus de l'abdomen lors du repos, mais l'apparition des deux mécanismes n'a peut-être pas été simultanée. Actuellement, certains groupes, en particulier tous les papillons, ont perdu secondairement cette capacité à se couvrir la surface du dos avec leurs ailes.
Chez les néoptères les plus primitifs, les deux paires d'ailes peuvent encore être indépendantes, mais chez les néoptères les plus évolués et possédant deux paires d'ailes fonctionnelles, celles-ci sont liées mécaniquement de diverses façons possibles, et fonctionnent de manière synchrone comme une seule paire.
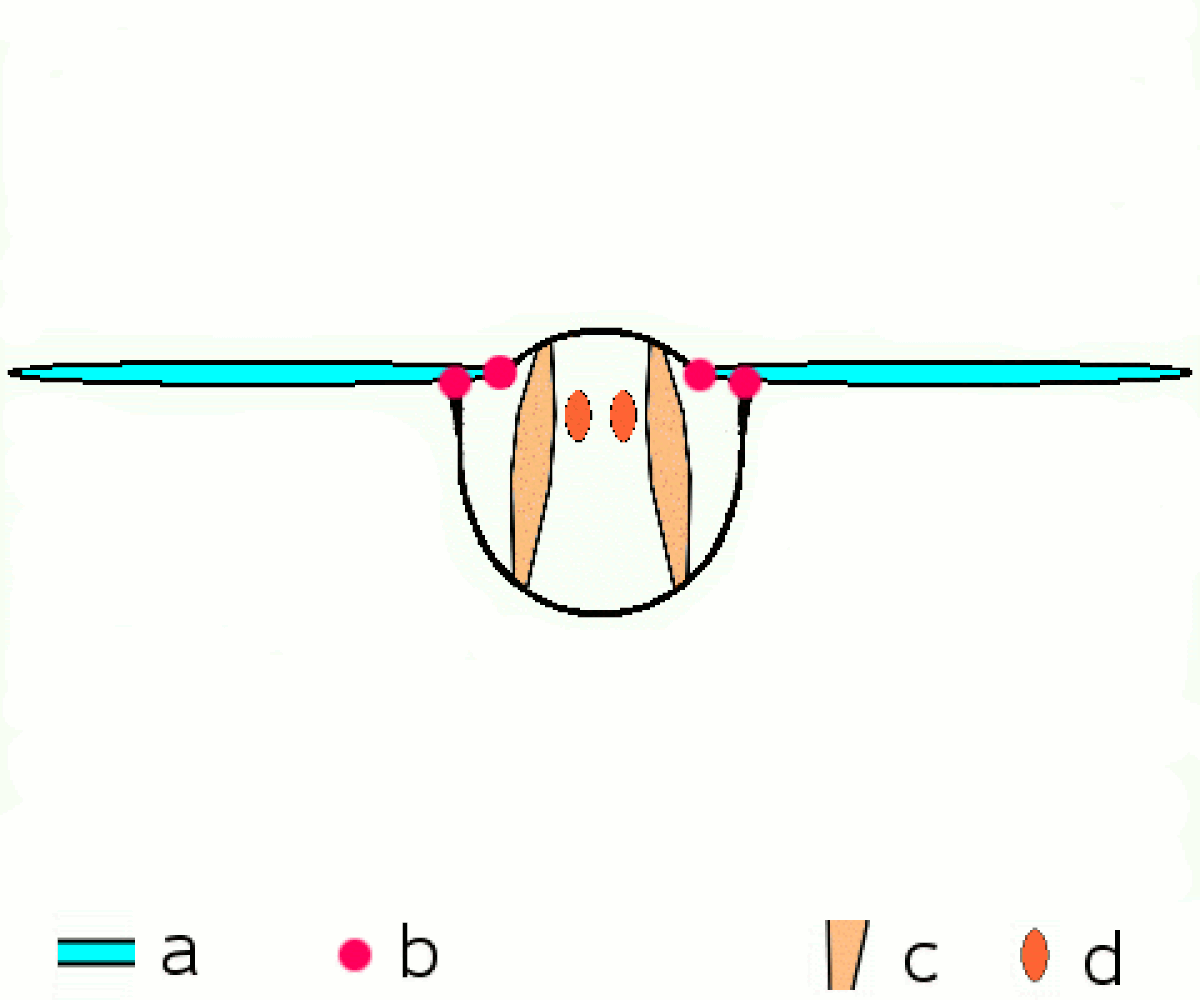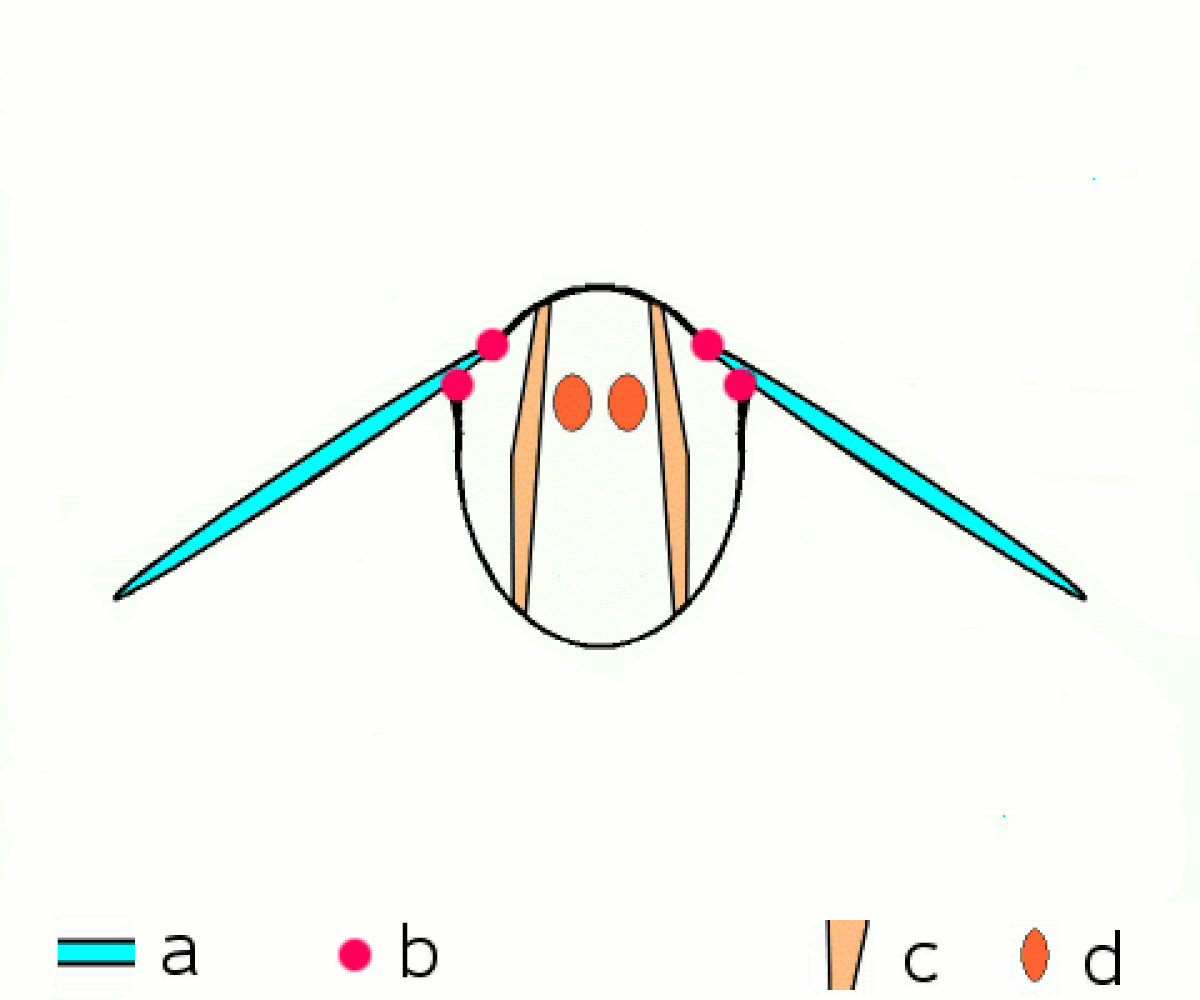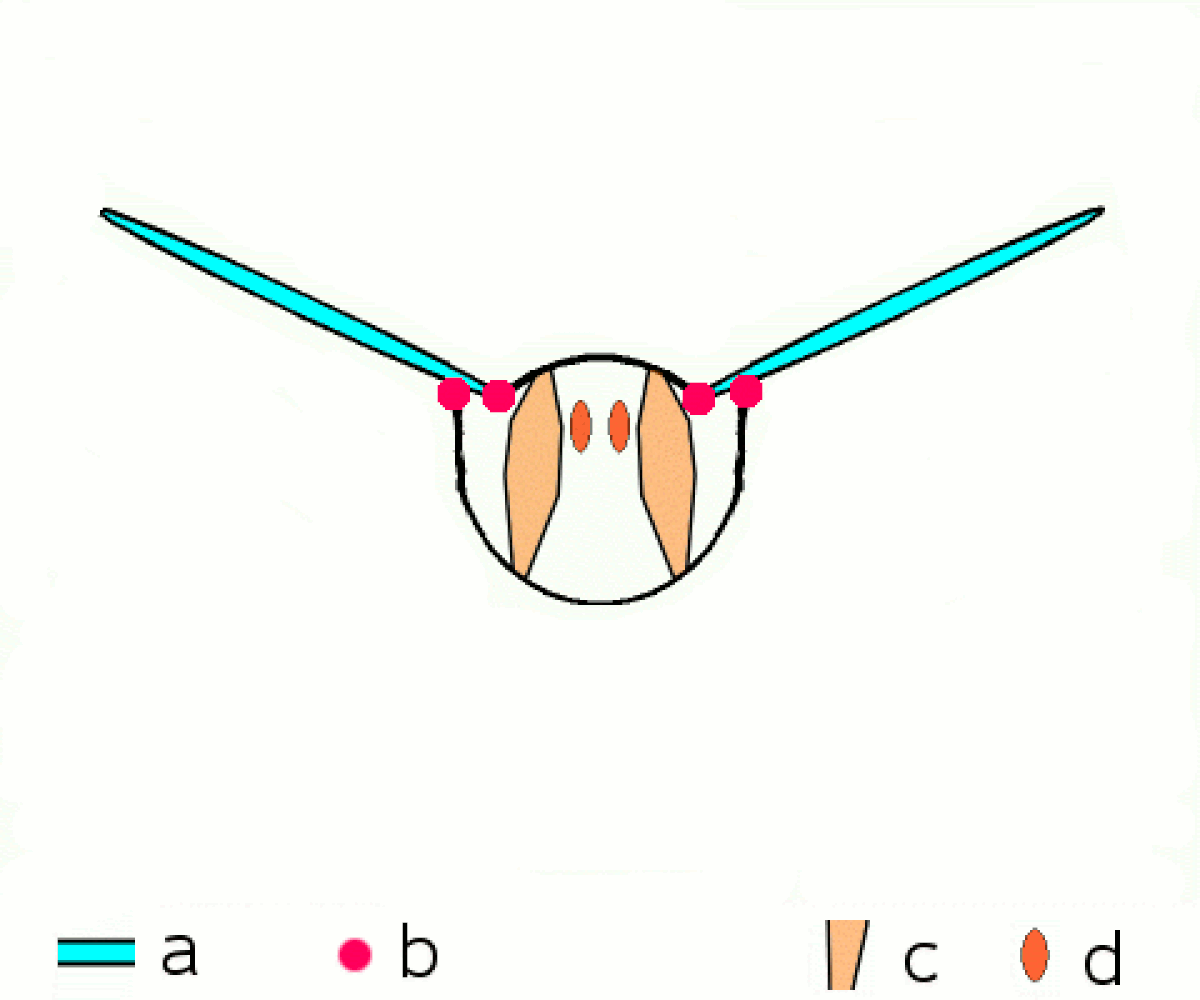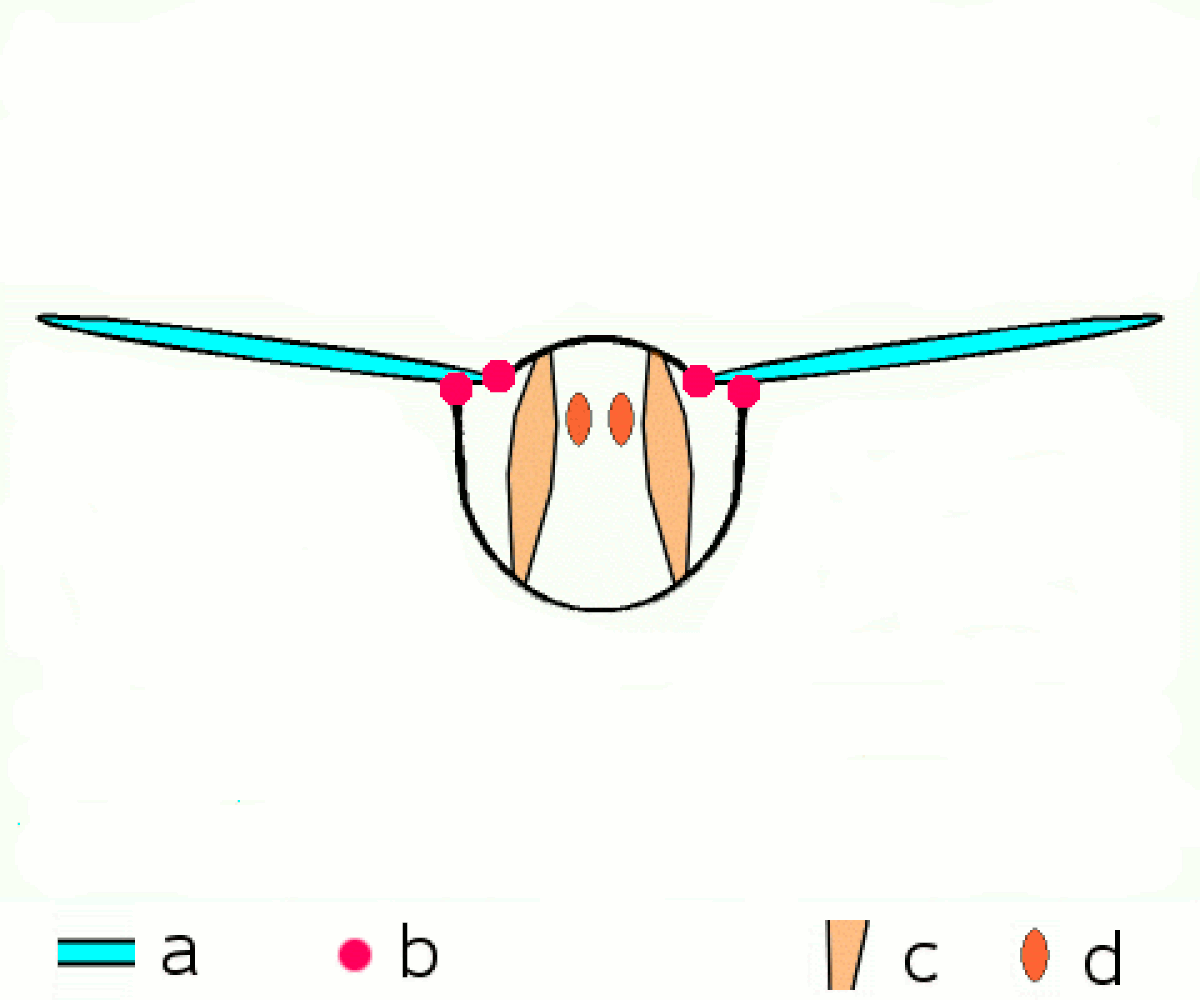
En revanche tous les néoptères partagent le même principe de transmission du mouvement aux ailes. Les muscles moteurs du vol, sont situés dans le thorax, y sont attachés et le déforment, et ce sont les déformations du thorax qui provoquent le mouvement des extensions de l'exosquelette thoracique que sont les ailes.
Une série de muscles dorsaux longitudinaux compriment le thorax de l'avant vers l'arrière, provoquant une courbure vers le haut de la surface dorsale du thorax, le notum), et faisant s'abaisser les ailes. Une série de muscles tergosternaux tirent le notum vers le bas de nouveau, provoquant le mouvement vers le haut des ailes. Dans quelques groupes, le battement vers le bas est accompli seulement par le recul élastique du thorax quand les muscles tergosternaux sont relâchés.
Plusieurs petites sclérites situées à la base de l'aile et attachées à d'autres séries de muscles, permettent d'ajuster finement la courbure et l'amplitude des battements d'ailes.
Un des ultimes raffinements apparu chez certains des néoptères supérieurs, tels les coléoptères, les diptères, et les hyménoptères, est un système de contrôle musculo-nerveux où une seule impulsion nerveuse entraîne la contraction répétée d'une fibre musculaire. Ceci permet à la fréquence des battements d'ailes d'excéder le rythme auquel le système nerveux peut envoyer des impulsions. Cet type de muscle est appelée « muscle de vol asynchrone ».
Le mécanisme de vol indirect, et ses diverses évolutions, a doté de nombreux néoptères supérieurs de capacités aériennes sophistiquées : vol stationnaire, à reculons, etc. avec une finesse et un degré de contrôle auquel les insectes ayant des muscles directement attachés aux ailes ne pouvaient prétendre.
Chez les diptères comme chez les coléoptères, le vol s'effectue à l'aide d'une seule paire d'aile. Chez les coléoptères, il s'agit de la seconde paire, la première étant transformée en élytres. Chez les diptères, c'est la première paire qui sert au vol, la seconde s'étant transformée en « haltères », qui sont de minuscules petites massues dont le mouvement sert à la mouche de gyroscope pour stabiliser son vol.
Mécanisme de vol direct

Chez les paléoptères, ordre de classification traditionnelle qui comprend les éphémères et les odonates, les muscles alaires, au contraire de la plupart des autres insectes, s'insèrent directement à la base des ailes. L'articulation est faite de telle façon qu'un petit mouvement vers le bas de la base de l'aile soulève le reste de l'aile vers le haut, à la manière d'une rame que l'on remonte en l'air.
Les ailes postérieures des éphémères sont réduites ou même quelquefois absentes et jouent un rôle mineur dans le vol, malhabile et disgracieux.
En revanche, chez les odonates (les demoiselles et les libellules), la paire d'ailes antérieures et la paire d'ailes postérieures sont semblables par la forme et par la taille. Elles opèrent indépendamment, ce qui permet un très fin degré de contrôle de la mobilité et une capacité, inégalée par les autres insectes volants, à pouvoir changer soudainement de direction et de vitesse, notamment pour chasser. La supériorité technique de ces prédateurs sur leurs proies pourrait expliquer la longévité de ce groupe biologique qui existait déjà bien avant l'apparition des dinosaures.
De plus les demoiselles peuvent tenir un vol stationnaire. Les libellules, avec des mécanismes de vol plus primitifs, n'y parviennent qu'imparfaitement et à condition de s'aider des courants d'air.