Histoire évolutive des cétacés - Définition
La liste des auteurs de cet article est disponible ici.
Protocetus

Présents il y a 45 Ma, les protocetidae forment une famille abondante et hétérogène dont on trouve des fossiles sur tous les continents. Le plus connu des genres étant celui des Rodhocetus. Ils disposaient de pattes plutôt adaptées à la nage mais leur laissant peut-être la possibilité de se déplacer sur le sol. On ne sait pas s'ils disposaient d'une nageoire caudale (horizontale comme les cétacés modernes).
Les protocétidés connus avaient de solides membres antérieurs et postérieurs, qui pouvaient soutenir le corps à terre, et il est probable qu'ils avaient une vie amphibie. Il est clair cependant qu'ils étaient mieux adaptés que leurs ancêtres à un style de vie aquatique. Chez rodhocetus, par example, le sacrum, qui chez les mammifères terrestres provient de la fusion de cinq vertèbres, reliant le pelvis avec le reste de la colonne vertébrale pour former le bassin, était divisé en vertèbres indépendantes, le pelvis n'étant plus connecté à la colonne vertébrale que par une vertèbre. De plus, ses narines étaient à mi-chemin du museau, première étape vers la position au sommet du crâne chez les cétacés modernes.

L'hypothèse d'une vie amphibie est soutenue par la découverte d'une femelle pleine de Maiacetus, chez laquelle le fœtus avait la position pour être mis bas par la tête, ce qui suggère que maiacetus mettait bas à terre.
L'appartenance des ancêtres de ces cétacés primitifs aux ongulés est rappelée par des caractères comme la présence de sabots au bout des doigts de Rodhocetus.
Ambulocétidés et remingtonocétidés
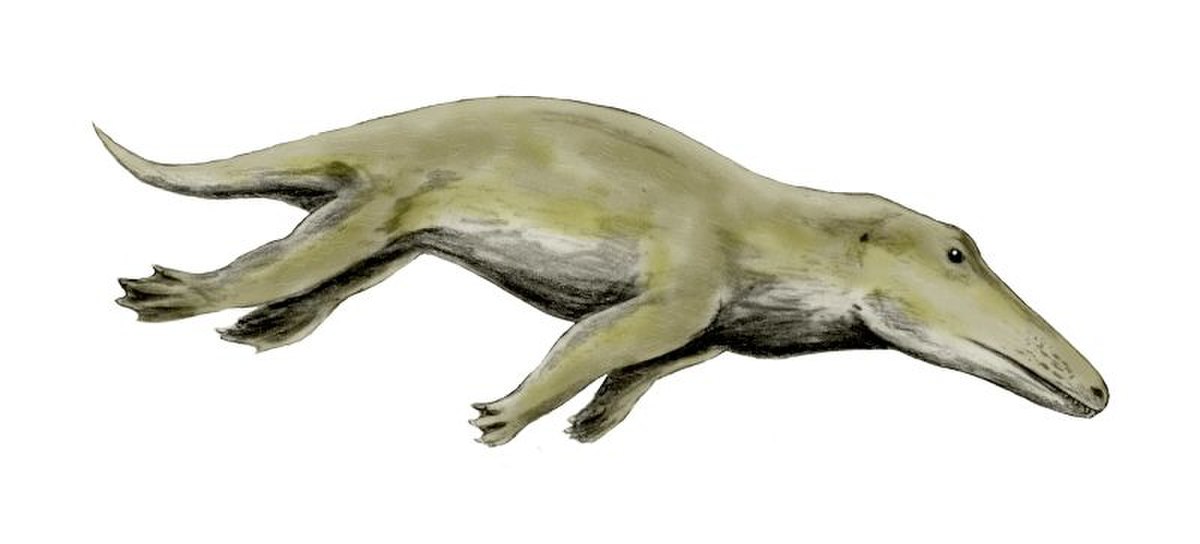
En 1994, on a fait la plus remarquable découverte, toujours au Pakistan, celle du fossile d'Ambulocetus clairement plus récent que ceux de Pakicetus. C'est un mammifère d'environ 3 m de long et aux pattes orientées vers l'extérieur, munies de petits sabots. Quoique clairement amphibie, il devait nager en ondulant l'arrière verticalement comme les loutres, les phoques et les cétacés, et être mal à l'aise sur la terre ferme. Alors que les Pakicetidae devaient vivre en eau douce, d'après certains travaux sur la composition isotopique des os, les Ambulocetus vivaient dans une grande variété de salinités. On a fait l'hypothèse que les ambulocétidés chassaient comme les crocodiles, tapis dans les lagunes pour attraper les imprudents, proies sur les bords, ou poissons.

Des cousins plus petits de l’ambulocetus formaient la famille des remingtonocétidés, qui possédaient des museaux plus longs qu’Ambulocetus, et étaient un peu mieux adaptés à la vie sous-marine. On pense qu'ils devaient avoir une vie analogue à celle des loutres de mer. L'examen des isotopes de leurs fossiles montre que tous les Remingtonocetidea trouvés vivaient dans un milieu marin. Dans les deux familles, les narines se trouvaient à l'extrémité du museau, comme chez les mammifères terrestres.
Les débuts de l'écholocation

Les crânes de squalodon sont les plus anciens à montrer la première apparition probable d'écholocation. Ce cétacé vivait de l'oligocène moyen au miocène moyen soit il y a de 33 à 14 Ma. Le squalodon montre plusieurs similitudes avc les odontocètes : le crâne est bien comprimé, le museau télescopé en-dehors (une caractéristique des odontocètes modernes), ce qui lui donnait une apparence de cétacé à dents. Ces animaux se font un panorama de leur environnement par écholocation sur le principe du sonar, c'est à dire en émettant une série de clics à diverses fréquences. Ces impulsions sonores sont émises par leur front en forme de melon, réfléchies par les objets, et recueillies, pour les hautes fréquences, par la mâchoire inférieure, d'où elles se propagent vers l'oreille.
Cependant il est improbable que ce soient des ancêtres des dauphins modernes.
Les premiers dauphins

Au début du Miocène de nombreuses espèces d'odontocètes ressemblant aux dauphins se sont développées comme les kentriodons et hadrodelphis. La plupart sont maintenant éteintes. On les classe dans les Kentriodontidae et ils ont la particularité d'avoir des crânes symétriques. Ils pourraient s'être déjà organisés en troupeaux. Ils consommaient des petits poissons et leur système d'écholocation devait être parfaitement opérationnel. La morphologie et la distribution de ces fossiles sont très semblables aux odontocètes modernes, qui pourraient y trouver leurs ancêtres. Les Odobenocetops sont des exemples de Delphinoidea du Pliocène.
Premières baleines à fanons (mysticètes)
On compte actuellement parmi les mysticètes 4 familles qui pourraient avoir des origines différentes chez les cétothères :
Tous les mysticètes modernes sont de grands cétacés à fanons, et se nourrissent en filtrant, bien que la manière exacte d'utiliser les fanons diffère entre les espèces (consommation par goulées chez les balénoptéridés, par écumage chez les baleines franches, et par aspiration du fond pour les baleines grises). Les premiers représentants de certains groupes modernes apparaissent au miocène moyen. Ceci peut avoir été dû à des changements environnementaux et physiques de l'océan. Un changement à grande échelle des courants et températures de l'océan peut avoir déclenché la radiation des mysticètes modernes, en éliminant les formes archaïques. En général, on pense que les quatre familles modernes de mysticètes ont des origines séparées parmi les cétothères : les Balaenopteridae (dont les baleines à bosse et les rorquals), les Balaenidae (baleines franches), les Eschrichtiidae (baleines grises), les Neobalaenidae (baleines pygmées) ont tous des caractères dérivés actuellement inconnus chez les cétothères.
Évolution du squelette
Aujourd'hui, les membes postérieurs des cétacés sont internes, et réduits. Ils ne servent plus que comme supports pour les muscles des organes génitaux. Alors que les cétacés primitifs comme le Pakicetus avaient les orifices nasaux au bout du museau, ces orifices ont commencé à migrer vers le sommet du crâne. Chez Rodhocetus, ils ont déjà substantiellement amorcé cette migration nasale. Les narines des cétacés modernes ont atteint le sommet du crâne et se sont modifiés en évents, qui leur permettent d'effleurer la surface, expirer, inspirer et replonger avec facilité. Les oreilles ont commencé à se déplacer aussi vers l'intérieur, et déjà chez le Basilosaurus, l'oreille moyenne commençait à recevoir des vibrations de la mâchoire inférieure. Les odontocètes modernes utilisent leur melon, un amas de graisse jouant le rôle de lentille acoustique pour l'émission de sons destinés à l'écholocation. Et ils utilisent leur mâchoire inférieure pour capter les échos de haute fréquence et les focaliser sur l'oreille.
Atavismes ancestraux
Les cétacés développent toujours certaines caractéristiques ancestrales à l'état embryonnaire ou de nouveau-nés. Les baleines à fanons développent des bourgeons de dents à l'état embryonnaire, qui disparaîtront chez l'animal adulte. Les fœtus de certaines espèces portent des poils éphémères qui disparaissent plus tard. Les embryons des cétacés ont les fosses nasales situées à l'extrémité du museau, comme chez tout mammifère. Celles-ci migrent durant le développement embryonnaire vers l'arrière, pour se stabiliser au-dessus du crâne et former l'évent chez le nouveau-né. Certains spécimens adultes disposent de moignons de pattes arrière. Les embryons de nombreuses espèces en ont également. Ce phénomène est connu sous le nom d'atavisme.
Arbre phylogénétique
Les arguments développés ci-dessus conduisent à une classification phylogénétique plus probable, encore que comme nous l'avons vu, certains détails restent à clarifier.
On en suivra les développements dans l'article Classification phylogénétique des Cetartiodactyla, qui est régulièrement mis à jour. Noter qu'en cliquant sur le lien [Dérouler] de l'onglet « Arbre développé », on peut le développer dans tous ses détails, ce qui n'est pas envisageable dans le présent article.