Nepenthes rajah - Définition
La liste des auteurs de cet article est disponible ici.
Classification
| | ||||||||
| N. maxima | N. pilosa | N. clipeata | ||||||
| N. oblanceolata * | N. burbidgeae | N. truncata | ||||||
| N. veitchii | N. rajah | N. fusca | ||||||
| N. ephippiata | N. boschiana | N. stenophylla | ||||||
| N. klossii | N. mollis | N. lowii | ||||||
| * Maintenant considéré comme un synonyme de N. maxima. | ||||||||
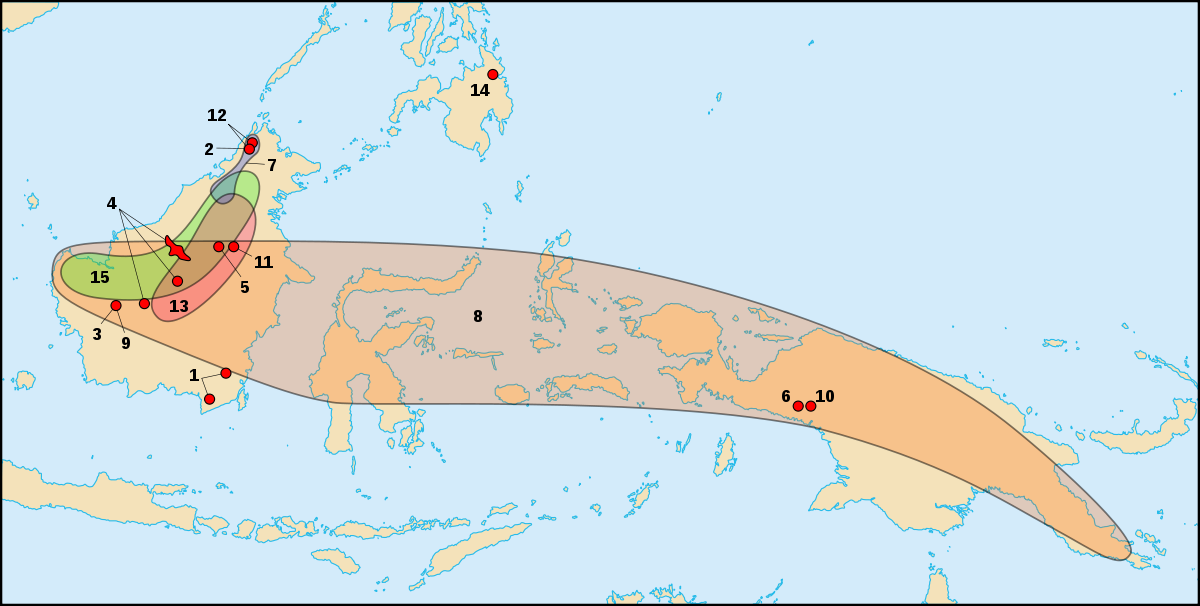
| ||||||||
| Distribution géographique du groupe des Regiae ; l'entrée 12 indique celle du Nepenthes rajah | ||||||||
Nepenthes rajah n'est généralement pas considéré comme étant proche d'une autre espèce, du fait de ses ascidies particulières et de la morphologie de ses feuilles. Toutefois, plusieurs tentatives ont été faites pour déduire des groupes logiques parmi le genre des Nepenthes, et ceci dans le but de montrer les relations entre les taxons inférieurs au genre. Ces tentatives ont groupées N. rajah avec d'autres espèces, censées partager des caractéristiques.
Au XIXe siècle
Le genre des Nepenthes a été scindé pour la première fois en 1873, quand Hooker publie sa monographie à propos de ce genre. Hooker distingue le N. pervillei de tous les autres espèces, car les graines de cette espèce ne portent pas les appendices qu'on trouve chez tous les autres Nepenthes (quoique grandement réduits chez N. madagascariensis). Il place N. pervillei dans le sous-genre monotypique Anurosperma (du latin, anuro : avec des nervures et sperma : graines). Toutes les autres espèces sont placées dans un second sous-genre, Eunepenthes (du latin, eu : vrai, les vrais Nepenthes).
Une seconde tentative pour l'établissement d'une division logique au sein du genre fut faite en 1895 par Beck dans son « Monographische Skizze » (en allemand, Monographische : monographie, Skizze: croquis). Beck conserve les deux sous-genres créés par Hooker, mais divise les Eunepenthes en trois sous-groupes : les Retiferae, les Apruinosae et les Pruinosae. N. rajah appartenant alors aux Apruinosae (du latin, pluriel de apruinosa : non dépoli). La plupart des taxonomistes contemporains de Beck ne donnaient qu'une faible valeur taxonomique à son groupement, puisque celui-ci était basé sur des traits arbitraires et non utilisables en tant que base pour une classification.
Au XXe siècle
La taxonomie des Nepenthes est révisée une fois de plus en 1908 par Macfarlane dans sa propre monographie. Singulièrement, Macfarlane ne nomme pas les groupes qu'il distingue. Sa proposition est souvent considérée comme non naturelle.
En 1928, B. H. Danser publia une monographie, dans laquelle il divise les Nepenthes en six clades qui sont : Vulgatae, Montanae, Nobiles, Regiae, Insignes et Urceolatae. Danser place le N. rajah dans le clade des Regiae (du latin, rēgia : royal). Le clade des Regiae, tel que proposé par Danser, est détaillé dans le tableau de droite.
La majorité des espèces de ce clade sont de grandes plantes avec des feuilles pétiolées, un indumentum de poils grossiers brun rouge, une inflorescence en racème, et des ascidies supérieures en forme d'entonnoir. Toutes sont pourvues d'un appendice caractéristique sur la surface inférieure de l'opercule près de l'apex. À l'exception de N. lowii, les Regiae possèdent toutes un péristome étendu et fascié (marqué de côtes). De plus une grande majorité des espèces appartenant à ce clade sont endémiques à Bornéo. En se basant sur la compréhension actuelle du genre, le clade Regiae semble plutôt bien refléter les relations unissant ses membres, bien qu'il n'en soit pas de même pour les autres clades. En dépit de cela, la classification de Danser est indubitablement une grande avancé au regard des tentatives antérieures.
Le travail taxonomique de Danser (1928) est révisé par Harms en 1936. Harms divise les Nepenthes en trois sous-genres : Anurosperma Hooker.f. (1873), Eunepenthes Hooker.f. (1873) et Mesonepenthes Harms (1936) (du latin, meso: milieu, Nepenthes « moyens »). Les espèces de Nepenthes situées dans les sous-genres Anurosperma et Mesonepenthes diffèrent de celles situées parmi les Vulgatae, où Danser les avait placées. Harms inclut N. rajah dans le sous-genre Eunepenthes avec la grande majorité des Nepenthes ; Anurosperma est un sous-genre monotypique, tandis que Mesonepenthes contient seulement trois espèces. Il crée également un clade supplémentaire, les Distillatoriae (d'après N. distillatoria).
Morphologie glandulaire
En 1976, Kurata suggère que les glandes présentes dans les ascidies des Nepenthes sont particulières à chaque espèce et peuvent ainsi être utilisées pour distinguer des taxons proches ou même être utilisées comme base de la classification. Kurata étudies deux types de glandes : les glandes à nectar sur l'opercule et les glandes digestives à l'intérieur de l'ascidie. Il divise ces dernières selon trois zones : supérieure, moyenne et inférieure. (Voir en:Image:Nepenthes_rajah_glands.png). Bien que cette nouvelle approche apporte un éclairage supplémentaire sur les similarités entre certaines espèces, elle a depuis été largement abandonnée par les taxonomistes et botanistes spécialisées dans ce genre, en faveur d'une nomenclature classique basée sur une description des caractères morphologiques.
Analyse biochimique
Plus récemment, des analyses biochimiques ont servi comme moyen de détermination des relations unissant différentes espèces de Nepenthes au sein d'un clade. En 1975, David E. Fairbrothers et al. suggèrent les premiers a link between chemical properties and certain morphological groupings ⇔ l'existence d'une relation entre les propriétés chimiques et certains groupements sur une base morphologique. Ils se basent sur la théorie selon laquelle des espèces végétales similaires sur le plan morphologique produisent des molécules chimiques aux effets thérapeutiques similaires.
En 2002, une étude basée sur des criblages phytochimiques et des chromatographies analytiques a été conduite sur la présence de composés phénoliques et de leucoanthocyanines chez plusieurs hybrides naturelles et leurs espèces parentes présumées (N. rajah y compris) provenant de Sabah et Sarawak. Elle a été effectuée sur des feuilles provenant de neuf spécimens séchés en herbier. Huit bandes ont été distinguées, elle contenaient des acides phénoliques, des flavonols, des flavones, des leucoanthocyanines et des « flavonoïdes inconnus » (numérotés 1 et 3). Leur répartition chez l'hybride N. × alisaputrana et ses espèces parentes présumées N. rajah et N. burbidgeae sont représentées dans le tableau suivant. Un spécimen de N. × alisaputrana issu de bouturage (in vitro) a également été testé.
| Distribution de composés phénoliques et de leucoanthocyanines chez N. × alisaputrana, N. burbidgeae et N. rajah | |||||||||
| Taxon | | | | | | | | | Spécimen |
| N. × alisaputrana | | | | | | | | | J2442 |
| in vitro | | | | | | | | | |
| N. burbidgeae | | | | | | | | | J2484 |
| N. rajah | | | | | | | | | J2443 |
| Légende : 1: acide phénolique, 2: acide ellagique, 3: quercétine, 4: kaempférol, 5: lutéoline, 6: 'Flavonoïde inconnu 1', 7: 'Flavonoïde inconnu 3', 8: Cyanidine ± : bande très faible, + : faible bande, ++ : bande bien présente, 3+ : bande très présente, | |||||||||
Les acides phénoliques et ellagiques n'ont pas été détectés chez N. rajah, tandis que des concentrations très faibles de kaempférol ont été détectées. Les chromatogrammes de N. × alisaputrana ont montré la complémentarité de ses deux parents présumés.
La myricétine est absente chez tous les taxa étudiés. C'est en accord avec les résultats antérieurs obtenus par d'autres auteurs (Som en 1988 ; Jay et Lebreton, 1972). L'absence de ce composé, largement présent au sein de la biosphère, chez les Nepenthes étudiés pourraient offrir « additional diagnostic information » (des informations supplémentaires) pour ces six espèces.
Séquençage
Plusieurs protéines et acides nucléiques du N. rajah ont été partiellement ou complètement séquencés. Les voici :
- translocated tRNA-Lys (trnK) pseudogene (DQ007139)
- trnK gene & maturase K (matK) gene (AF315879)
- trnK gene & maturase K (matK) gene (AF315880)
- maturase K (AAK56010)
- maturase K (AAK56011)
Une espèce apparentée ?
Récemment, une nouvelle espèce remarquable de Nepenthes a été découverte aux Philippines par Andreas Wistuba. Surnommé temporairement N. sp. Palawan 1, ce népenthès ressemble fortement au N. rajah au niveau de la morphologie des ascidies et des feuilles. (Voir [1][2][3]). Du fait de la distance géographique séparant les deux espèces, il est peu probable qu'elles soient étroitement apparentées de quelque façon que ce soit. Ainsi, ce cas pourrait représenter un exemple d'évolution convergente, processus par lequel deux organismes non étroitement apparentés acquièrent indépendamment des caractéristiques similaires, évoluant dans des écosystèmes différents, mais comparables. La description formelle de N. sp. Palawan 1 est en cours.