Anatomie de la barrière hémato-encéphalique - Définition
La liste des auteurs de cet article est disponible ici.
Domaines du cerveau sans barrière hémato-encéphalique
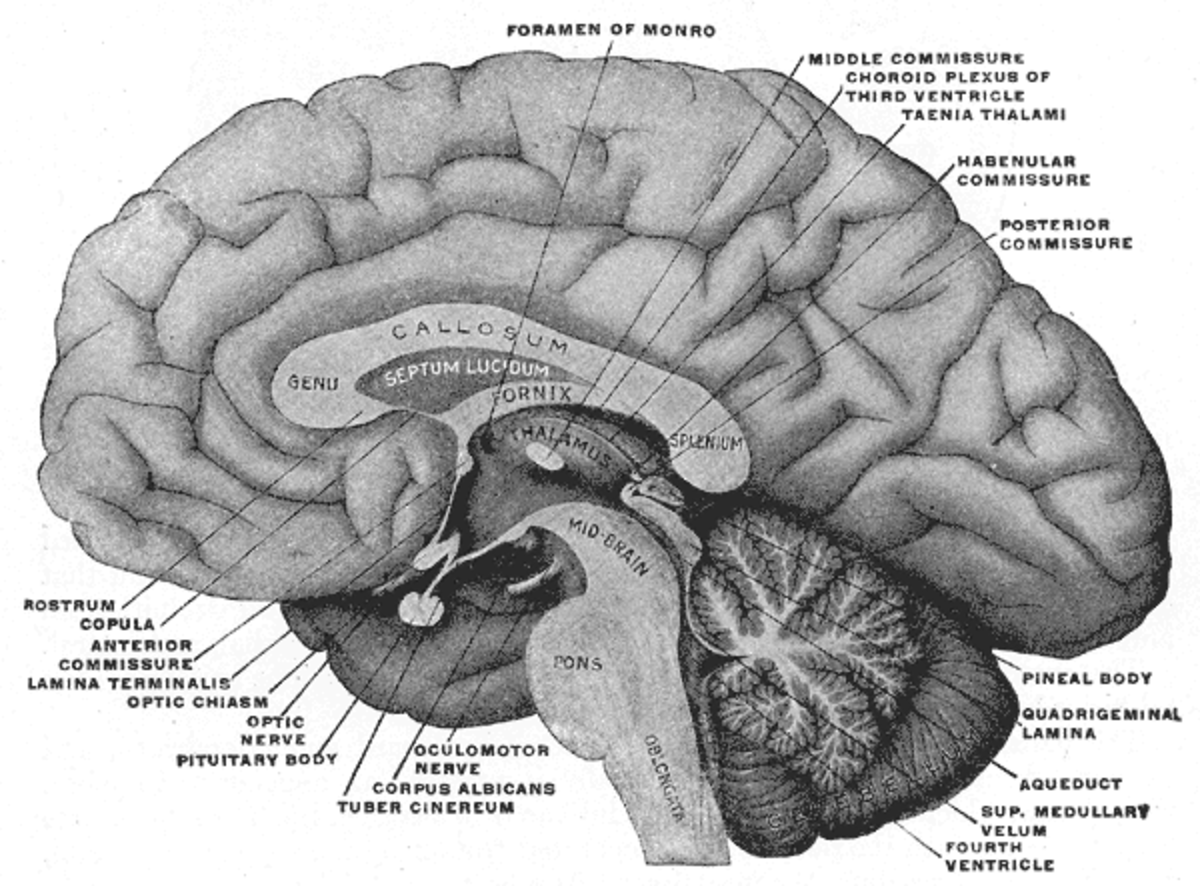

Tous les capillaires du cerveau ne font pas partie de la barrière hémato-encéphalique. Par exemple, les régions cérébrales qui déversent dans le sang des substances neuroendocrines (hormones), ou encore qui ont une fonction sensorielle – par exemple des zones réceptrices de certaines hormones peptidiques – ne peuvent pas avoir de barrière hémato-céphalique. C'est le cas pour six organes circumventriculaires. L'épiphyse, l'éminence médiane, la neurohypophyse, l'area postrema, l'organe vasculaire de la lame terminale (OVLT) et l'organe subfornical présentent des capillaires fenestrés. Par exemple, c'est ainsi que les neurones de l'area postrema peuvent détecter des substances toxiques, et déclencher un besoin de vomir dans le centre du vomissement. L'organe subcommissural est le seul des organes circumventriculaires qui présente une barrière hémato-encéphalique. Pour protéger les tissus cérébraux avoisinants, les organes circumventriculaires sont entourés de tanycytes. Ce sont des épendymocytes spéciaux, avec des jonctions serrées très étanches.
Les astrocytes
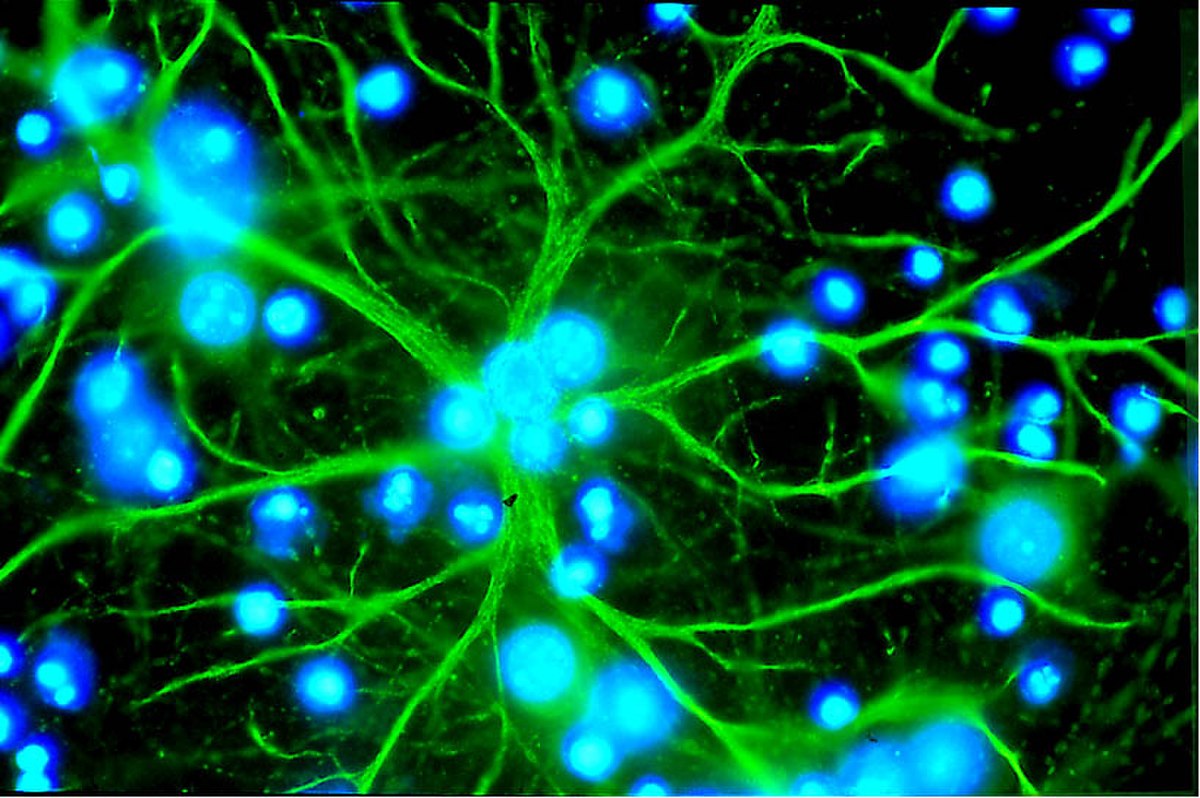
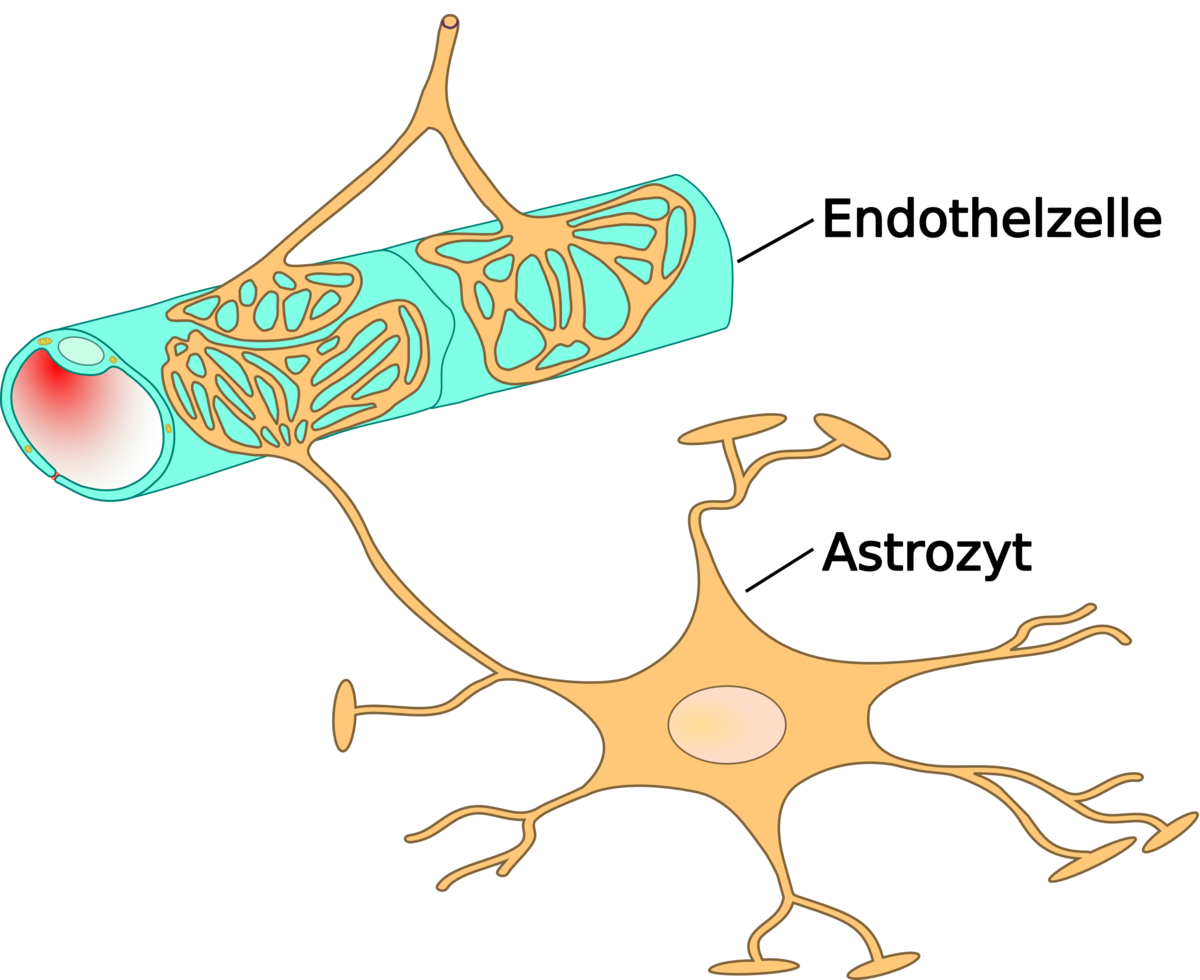
Les astrocytes sont des cellules en étoile de la famille des macroglies, significativement plus grandes que les péricytes. Ils font partie du système nerveux central, et sont capable de division encore après la naissance. Chez les vertébrés supérieurs, ils n'ont aucune fonction directe de barrière, bien qu'ils couvrent les capillaires du cerveau à 99% avec leurs pieds. Ils sont cependant en interaction immédiate avec les cellules endothéliales. Les astrocytes induisent sur les cellules endothéliales des vaisseaux cérébraux la fonction de barrière hémato-encéphalique. Ceci a été démontré par des expériences de transplantation : des capillaires cérébraux transplantés dans des organes périphériques se sont comportés comme les capillaires « normaux » voisins, en développant par exemple des fenestrages. Inversement, des capillaires périphériques transplantés dans le système nerveux central prennent l'aspect local, avec des jonctions serrées. Cette influence des astrocytes sur l'aspect des cellules endothéliales se montre aussi dans des expériences in vitro : dans des cultures simultanées d'astrocytes et de cellules endothéliales, les endothéliums se montrent plus serrés que dans des cultures pures de cellules endothéliales.
Les astrocytes répandent un ensemble d'hormones, qui peuvent moduler la perméabilité de l'endothélium en l'espace de secondes ou de minutes. En sens inverse, les cellules endothéliales secrètent du facteur inhibiteur de la leucémie (LIF), une cytokine du groupe des interleukine 6, qui induit la différenciation des astrocytes. La distance entre les pieds d'astrocytes et les cellules endothéliales et péricytes n'est que de 20 nm.
La fonction principale des astrocytes est cependant l'alimentation des neurones, ainsi que la régulation de la concentration ionique extracellulaire. Une grande partie du cholestérol présent dans le cerveau est produit par les astrocytes. Le cholestérol ne peut pas traverser la barrière hémato-encéphalique, et il faut donc le synthétiser sur place. On trouve dans le cerveau environ 25% du cholestérol du corps entier ; essentiellement dans la myéline qui gaine les axones des neurones.
Les pieds des astrocytes forment un contact en filigrane de rosettes sur les cellules endothéliales. Cet arrangement est important pour l'influence réciproque et la communication entre les deux types de cellules. C'est ainsi qu'une diffusion libre est rendue possible entre les cellules endothéliales et le parenchyme cérébral.
Les maladies qui concernent directement ou indirectement les astrocytes, comme la maladie d'Alzheimer ou les astrocytomes, peuvent altérer substantiellement les fonctions de la barrière hémato-encéphalique, en raison de leur interaction étroite.

















































