Anatomie de la barrière hémato-encéphalique - Définition
La liste des auteurs de cet article est disponible ici.
L'endothélium
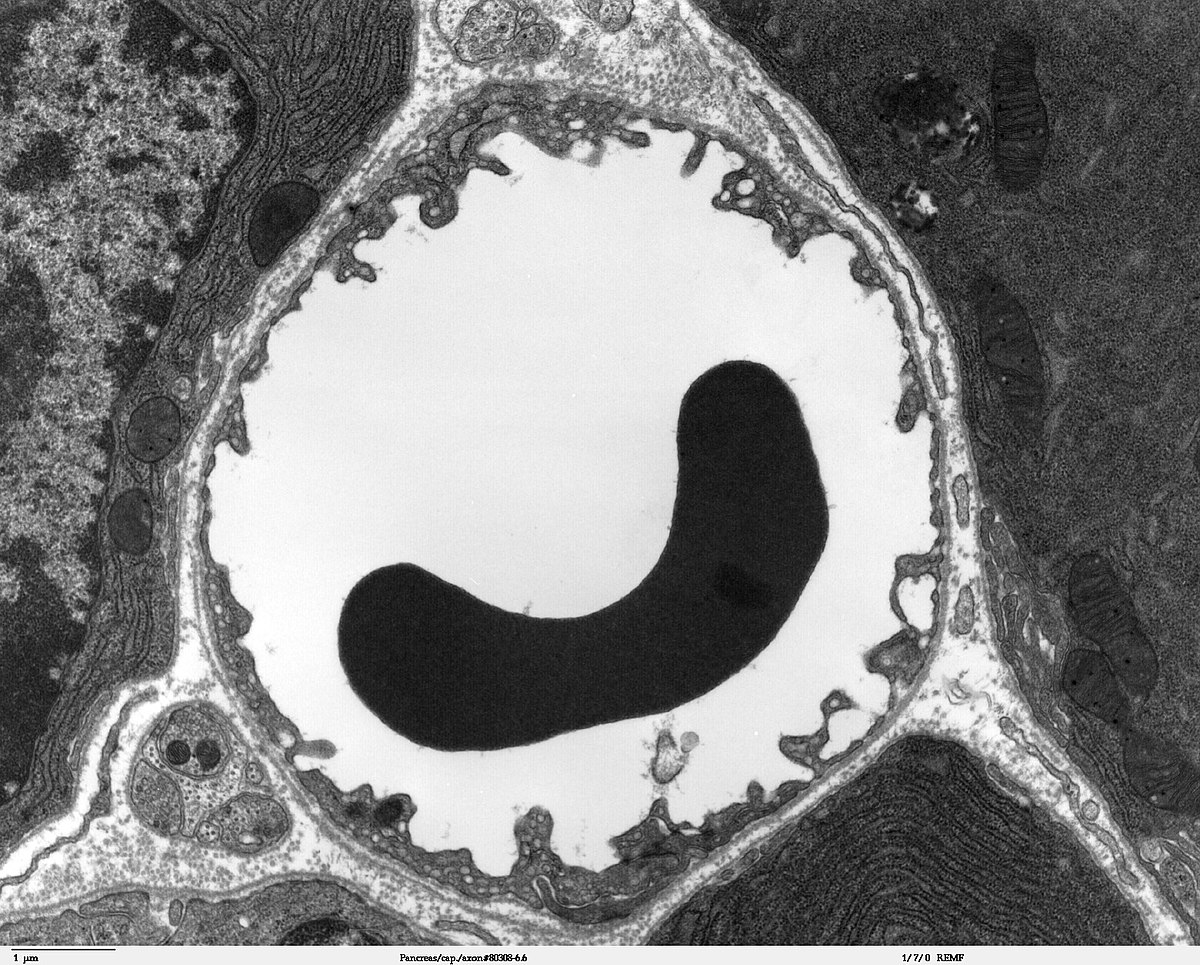
Les capillaires sont tapissés – comme les vaisseaux périphériques – de cellules endothéliales. L'endothélium des capillaires périphériques possède, pour l'échange de l'eau et des matières qui y sont dissoutes ou en suspension entre le sang et le liquide intercellulaire du tissu environnant, des ouvertures (fenêtres) d'environ 50 nm de diamètre et des fentes entre cellules de 0,1 à 1 µm de large. Entre les cellules endothéliales du cerveau, il n'y a au contraire aucun fenestrage, ni fente intercellulaire, ce pourquoi on parle aussi d'un « endothélium continu ». La membrane cellulaire du côté de l’intérieur du capillaire (« luminale ») diffère considérablement du point de vue de la composition en protéines de celle de la membrane du côté du tissu interstitiel (« apicale »).
La quantité de vésicules de pinocytose, qui permettent l'endocytose de substances dissoutes, est très faible dans l'endothélium du cerveau.
Par contrase, le nombre des mitochondries est environ de 5 à 10 fois plus élevé que dans les capillaires périphériques. Ceci indique le besoin élevé en énergie de l'endothélium – entre autres pour les processus de transport actif – et un métabolisme élevé. La barrière hémato-encéphalique se montre donc une barrière non seulement physique, mais encore métabolique, ou plutôt enzymatique. Dans la membrane cellulaire de l'endothélium, on trouve une série d'enzymes en quantité particulièrement élevée, comme par exemple dans les cellules du parenchyme. Ces enzymes comprennent notamment la γ-Glutamyltransférase, la phosphatase alcaline et la glucose-6-phosphatase. Les enzymes métaboliques comme la catéchol-0-méthyltransférase, la monoaminooxydase ou le cytochrome P450 sont également actives en relativement haute concentration dans l'endothélium De cette manière, beaucoup de substances sont métabolisées avant ou pendant leur transport intracellulaire. Les cellules endothéliales sont minces, de 0,3 jusqu'au plus 0,5 µm. Par comparaison, les entérocytes, cellules épithéliales de l'intestin, sont bien plus épaisses, de 17 à 30 µm.. Le rapport du cholestérol aux phospholipides se situe, comme pour les autres cellules endothéliales à 0,7. Les processus de simple diffusion à travers la membrane cellulaire sont donc aussi rapides au niveau de la barrière hémato-encéphalique que dans les autres épithéliums. Les cellules endothéliales présentent sur leurs membranes une quantité d'aquaporines pour la régulation de la quantité d'eau au sein du cerveau. Ces canaux spécialisés permettent une libre diffusion des molécules d'eau dans les deux sens.
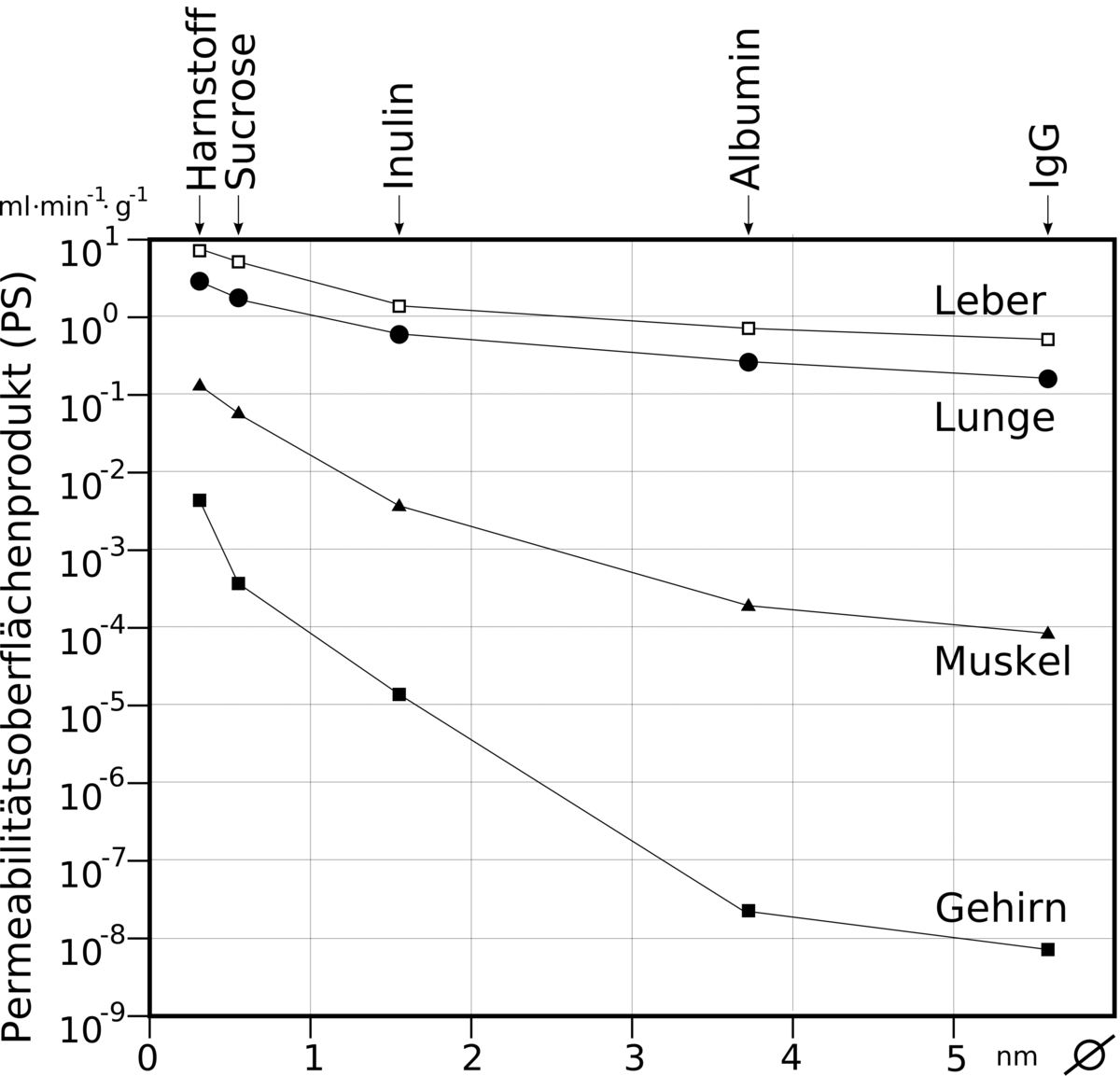
L'absence de fenestrages et le manque de vésicules pinocytotiques représentent l'aspect passif de la barrière hémato-encéphalique. Cet effet de barrière peut être quantifié par sa résistance électrique. Chez un rat adulte et sain, la résistance monte à environ 1 500 à 2 000 Ω⋅cm². Par opposition, la valeur pour des capillaires de tissu musculaire est d'environ 30 Ω⋅cm².
Les systèmes de transport actifs de l’endothélium décrits plus loin en plus grand détail constituent la partie active de la barrière hémato-encéphalique.
Les jonctions serrées
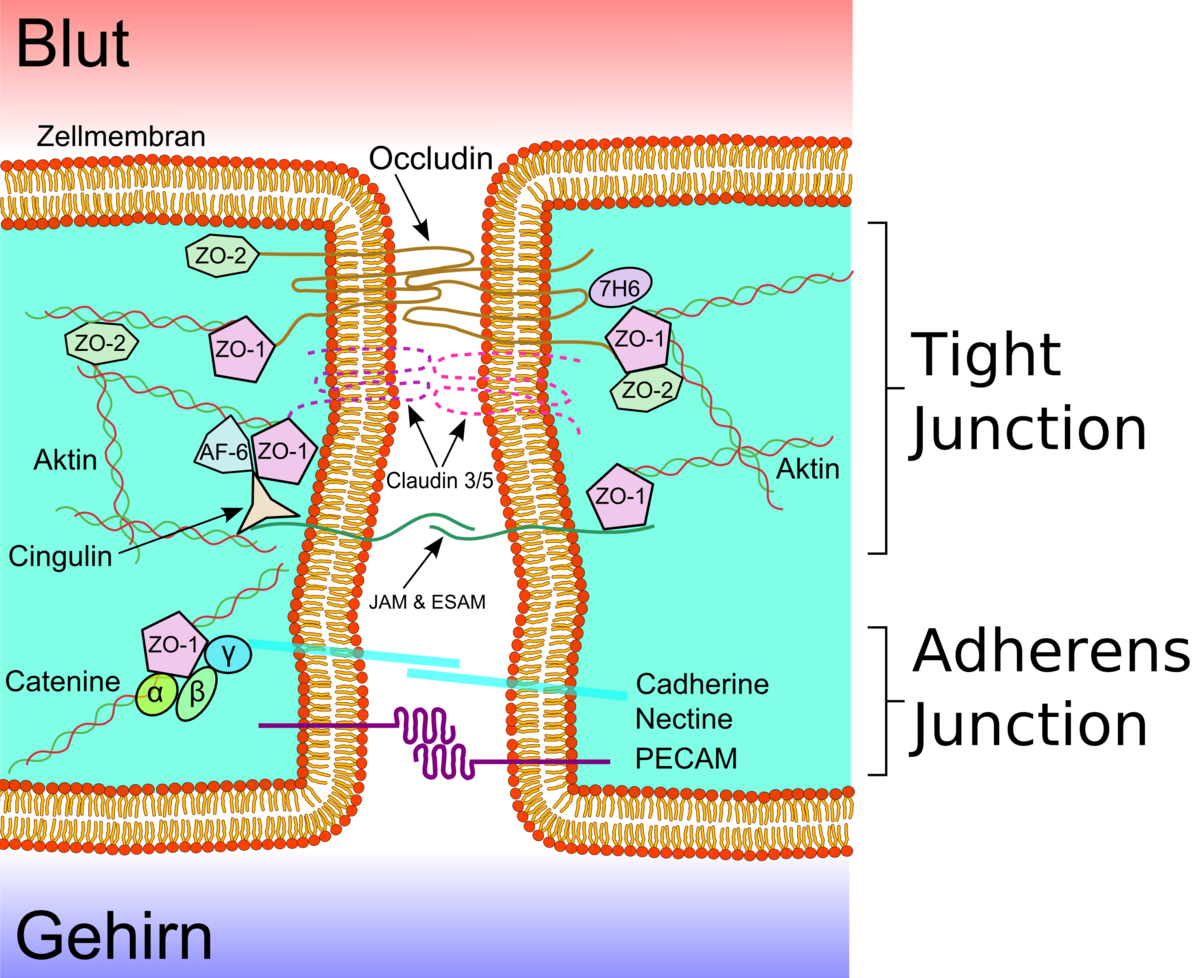

Les cellules endothéliales sont liées ensemble par de solides liaisons, appelées jonctions serrées. Ces jonctions contribuent substantiellement à la fonction de la barrière hémato-encéphalique. Elles ferment pratiquement complètement la voie de transport paracellulaire, c'est-à-dire l'échange de matières entre les membranes de deux cellules endothéliales voisines.
Plusieurs protéines membranaires comme l'occludine, diverses claudines et molécules d'adhésion de jonction (JAM sous leur sigle anglophone) relient les cellules endothéliales dans la région apicale des parois cellulaires latérales en bandes de protéines qui font un tour complet, et se connectent en jonctions serrées, qui interdisent largement le transport de matières entre les membranes cellulaires endothéliales Les liaisons protéine-protéine travaillent comme des sécurités en parallèle, c'est-à-dire perdent leur efficacité dès que l'une cède.
Les cellules endothéliales de la barrière hémato-encéphalique expriment surtout deux des 24 claudines connues, la C-5 et la C-12. La C-5 est considérée comme la plus importante des molécules d'adhésion de cellules dans la barrière hémato-encéphalique. La désactivation du gène CLDN5 qui code pour la claudine-5, conduit chez les souris ainsi modifiées à ce que la barrière hémato-encéphalique devient perméable aux molécules de masse molaire inférieure à 800 g/mol. Les animaux ainsi génétiquement modifiés meurent dans les quelques heures qui suivent leur naissance.
Par fixation, l'endothélium montre ses deux faces très différentes : la face luminale, du côté du sang, et la face abluminale, du côté du cerveau.
La lame basale
Les cellules endothéliales sont complètement entourées par une couche protéique, la lame basale. Cette double couche épaisse de 40 à 50 nm n'est visible qu'au microscope électronique. Elle est constituée essentiellement de collagène de type IV, de protéoglycanes-héparan-sulfate, de laminine, de fibronectine et autres protéines de la matrice extracellulaire. Cette lame basale touche la membrane plasmatique des pieds astrocytaires.