Synthèse des protéines - Définition
La liste des auteurs de cet article est disponible ici.
Introduction
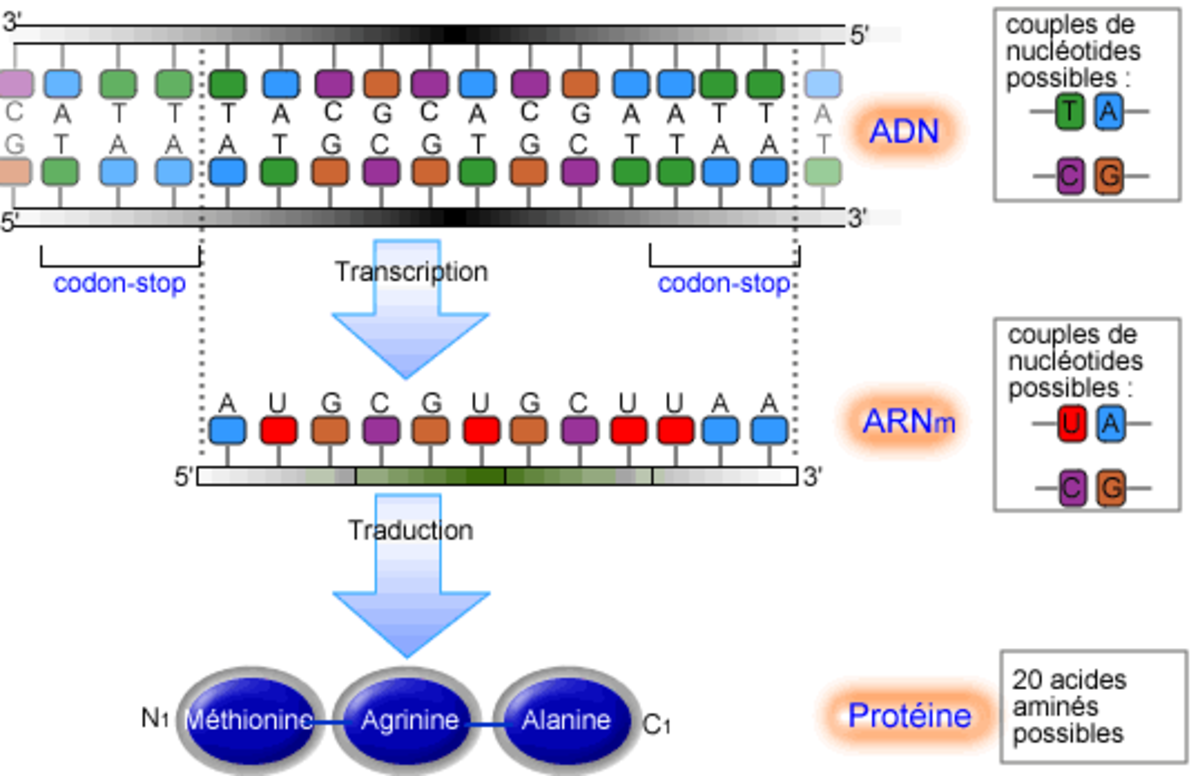
La synthèse des protéines est l'acte par lequel une cellule assemble une chaîne protéique en combinant des acides aminés isolés présents dans son cytoplasme, guidé par l'information contenue dans l'ADN. Elle se déroule en deux étapes au moins : la transcription de l'ADN en ARN messager et la traduction de l'ARN messager en une protéine.
Chez les eucaryotes, il existe une étape intermédiaire, la maturation de l'ARN prémessager, qui se passe dans le noyau. L'ARN prémessager subit l'ajout d'une coiffe de 7-méthylguanosine triphosphate à l'extrémité 5' et d'une queue poly(A) (50 à 250 nucléotides d'adénine) à l'extrémité 3'. Par la suite, l'ARN prémessager subit une excision de ses introns (les parties du gène qui ne codent pas un polypeptide) et l'épissage de ses exons (les brins codant). L'ARN prémessager est maintenant à maturité et prend le nom d'ARN messager. La transcription se déroule dans le noyau, la traduction, dans le réticulum endoplasmique. Une dernière étape de glycosylation (liaison covalente d'oses aux protéines) a lieu dans l'appareil de Golgi. Chez les procaryotes, les deux étapes ont lieu dans le cytoplasme et peuvent être simultanées, la traduction débutant alors que la transcription n'est pas encore achevée. Cette simultanéité donne lieu à un important type de régulation de la traduction.
Première approche : mécanisme global
Mise en évidence du mécanisme global de synthèse (pulse-chase)
Il est légitime de se demander comment les biologistes ont découvert la succession d'étapes qui mène à l'achèvement des protéines. La technique principale est celle du ou pulse-chasse, qui se déroule en quatre étapes principales.
On prélève à intervalles réguliers des cellules de ce nouveau milieu ; on dispose alors de deux techniques d'exploitation de cette expérience. La première est l'autoradiographie ; la seconde passe par une ultracentrifugation et donne une meilleure précision dans les résultats. On prépare une coupe de la cellule, par fixation puis découpage au microtome. Sur la plaquette obtenue, on dépose un film photographique contenant des grains d'argent, et on laisse le tout reposer quelques semaines à l'obscurité. Les électrons issus de la désintégration des noyaux radioactifs assimilés par la cellule réduisent les ions Ag+ en grains noirs d'argent, donnant ainsi une « photographie » de la localisation de la radioactivité cellulaire. On peut ainsi retracer, par observation de lames minces à temps de « chasse » différents, le trajet cellulaire des protéines lors de leur synthèse. Cependant les électrons de la lame mince peuvent marquer la plaque photographique assez loin de leur zone d'émission (précision de l'ordre du demi-micromètre). Ainsi, on ne peut pas savoir par exemple, si la radioactivité dans un organite est à l'intérieur ou éventuellement juste à l'extérieur de ses parois. On centrifuge à haute vitesse chaque prélèvement de cellules du second milieu. On obtient ainsi, après plusieurs centrifugations successives à des accélérations croissantes, différentes fractions de cellule, classées suivant leur masse. On connaît la correspondance entre les divers organites (réticulum endoplasmique, appareil de Golgi, noyau) et les fractions après centrifugations. Ainsi, si on mesure la radioactivité de chaque fraction, on peut savoir dans quel organite les acides aminés marqués se trouvaient au moment du prélèvement. On en déduit des courbes de répartition de radioactivité en fonction du temps pour chaque compartiment cellulaire, ce qui permet de retrouver le trajet des acides aminés nouvellement assemblés en protéines.
Complémentarité des bases azotées
Il est important de savoir, avant de procéder, qu'il existe une complémentarité entre les bases azotées de l'ADN et de l'ARN. C'est cette complémentarité, due à des liaisons hydrogène, qui permet la réplication, la transcription et la traduction des acides nucléiques.
Il existe 5 bases azotées :
- Adénine (A)
- Guanine (G)
- Thymine (T) qu'on ne trouve que dans l'ADN
- Cytosine (C)
- Uracile (U) qu'on ne trouve que dans l'ARN
A et G sont des purines (2 cycles)
T, C et U sont des pyrimidines (1 cycle)
A et T sont complémentaires
G et C sont complémentaires
A et U sont complémentaires
La transcription et la traduction utilisent cette complémentarité lors de la création d'ARN et de l'approche de l'ARNt.

















































