Cervelet - Définition
La liste des auteurs de cet article est disponible ici.
Introduction
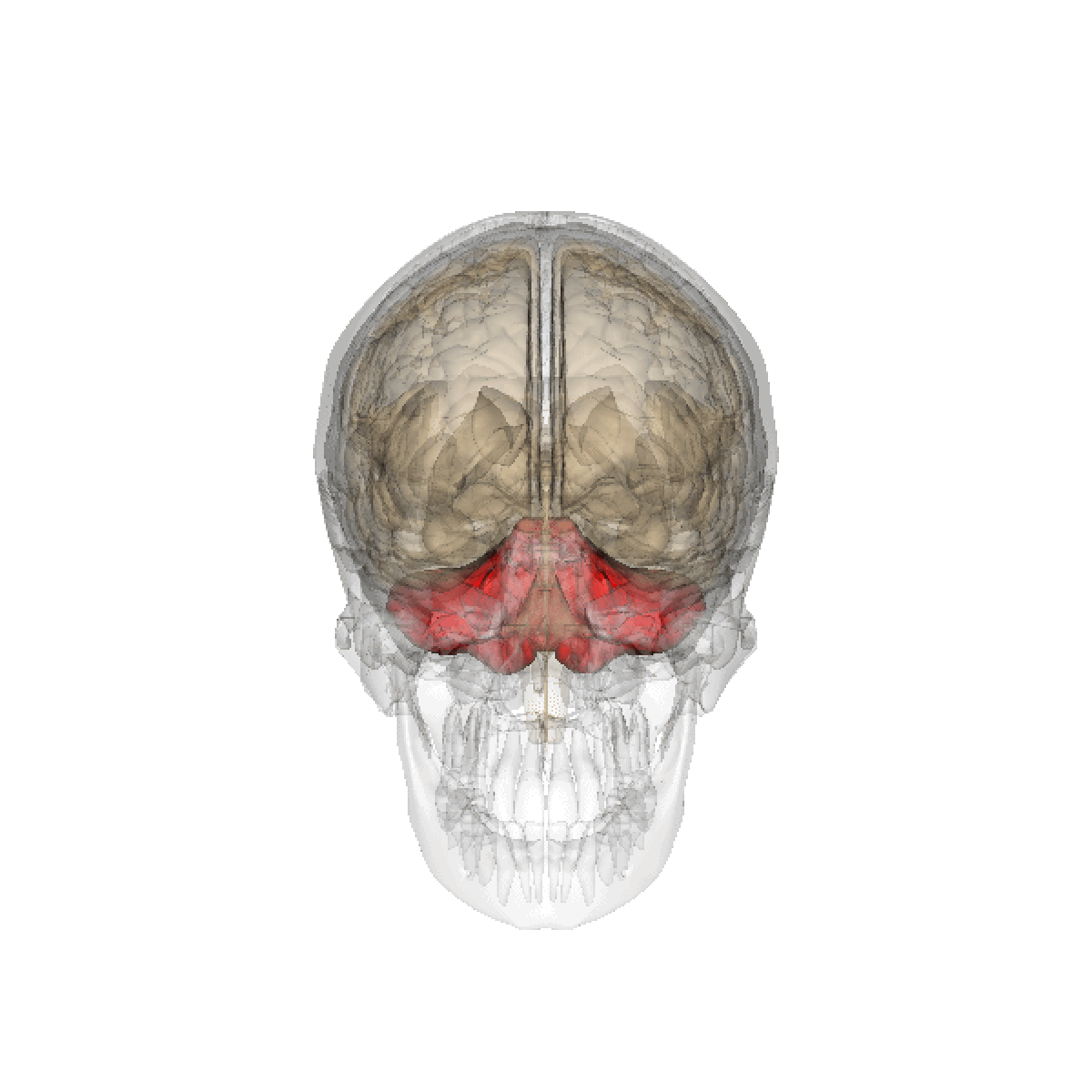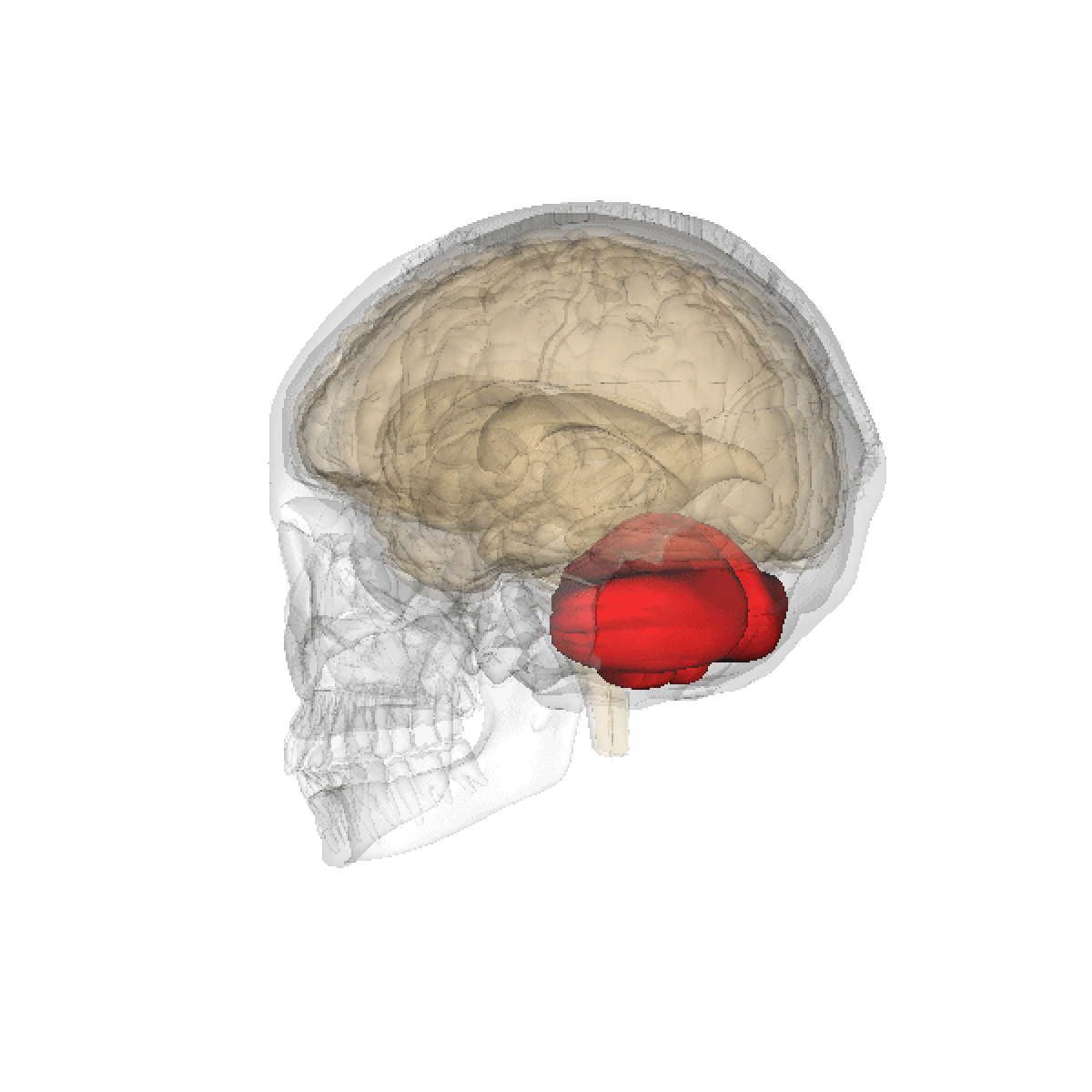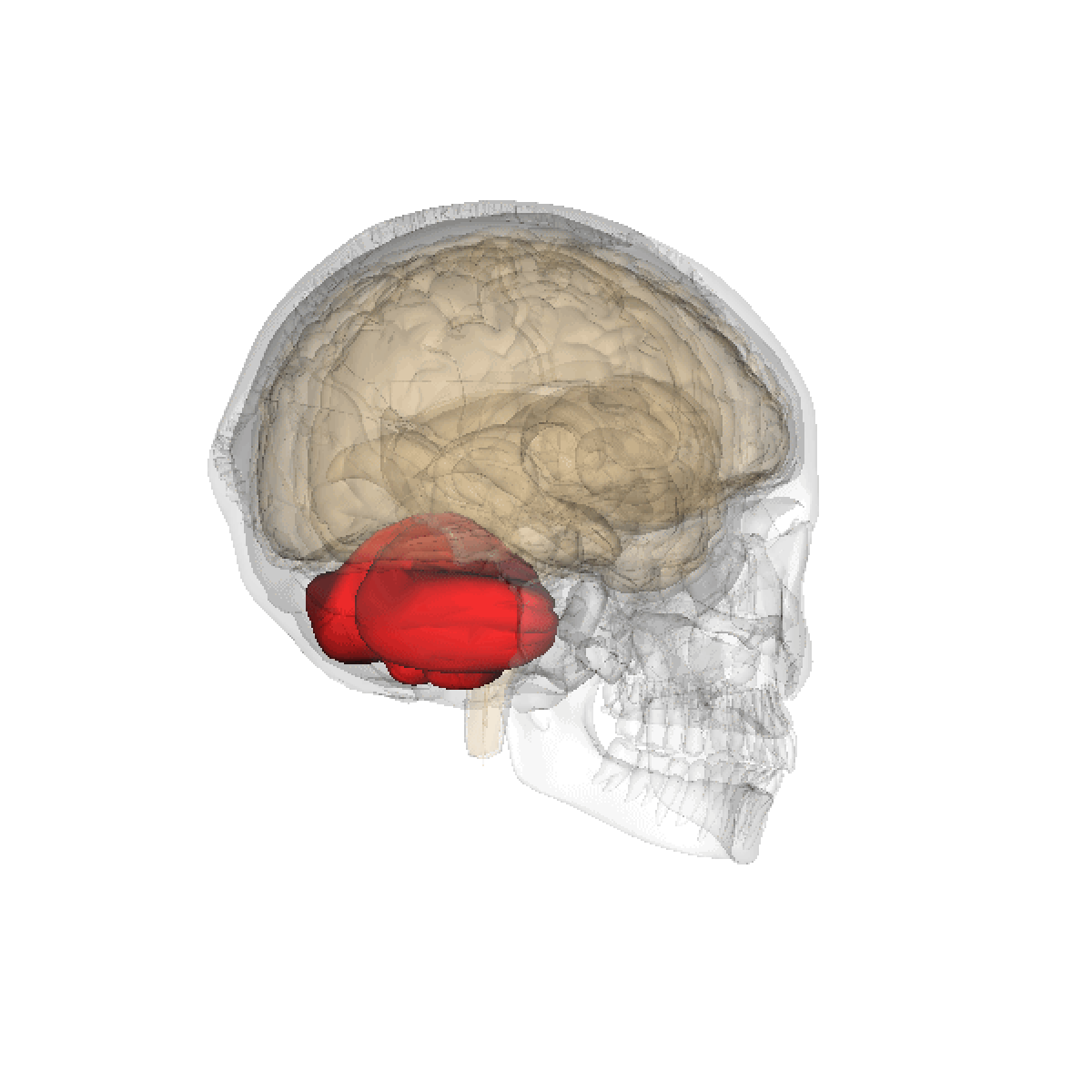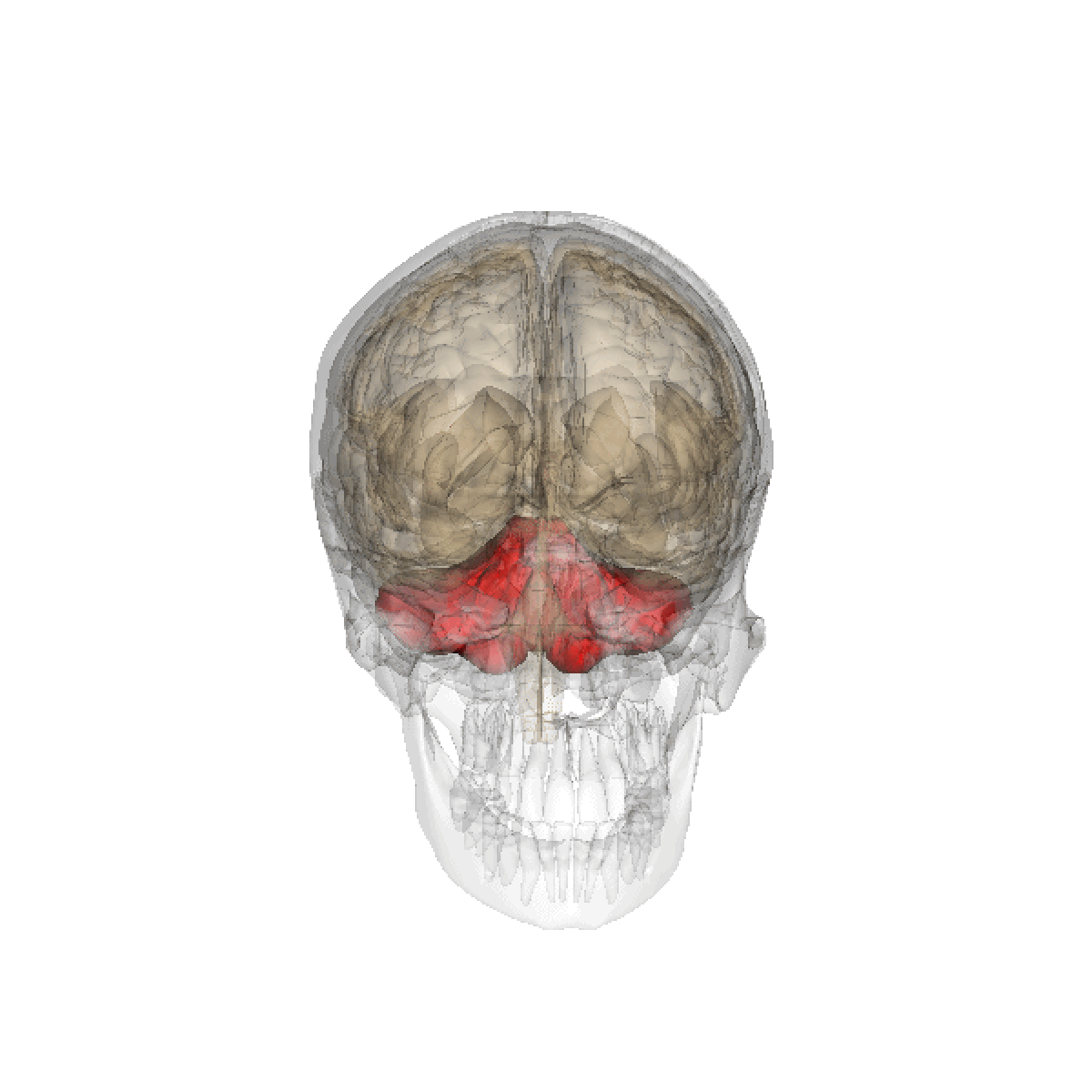

Le cervelet est une importante structure de l'encéphale (cortex trilaminaire), situé sous le cerveau proprement dit et derrière le tronc cérébral. Il n'occupe que 10% environ de la boîte crânienne mais comprend plus de 50% de ses neurones.
Depuis le XVIIIe siècle, différentes observations ont montré que le cervelet joue un rôle majeur dans la coordination des mouvements. Les lésions du cervelet ne provoquent pas de paralysie mais elles entrainent des difficultés lors de l'exécution et de l'apprentissage des mouvement complexes associées à des troubles de l'équilibre. Depuis la fin du XXe siècle, le cervelet est incriminé dans des fonctions non motrices: sensorielles, affectives et cognitives.
Au cours du développement embryonnaire, le cervelet se développe à partir du métencéphale, comme le pont de Varole, et du mésencéphale.
Généralités
Le cervelet est situé en dessous du lobe occipital du cerveau, et en arrière du tronc cérébral, en particulier, en arrière du bulbe et de la protubérance.
- Situation : dans la fosse postérieure du crâne, en arrière du tronc cérébral et sous la face ventrale du lobe occipital dont il est séparé par la tente du cervelet.
- Structure : cortex tri-laminaire se projetant sur des noyaux cérébelleux profonds.
- Origine embryologique : rhombencéphale et, partiellement, mésencéphale.
- Origine phylogénétique: ostracodermes (protochordés).
- Subdivisions phylogénétiques:
- archicerebellum à partir des poissons cartilagineux;
- paleocerebellum à partir des reptiles;
- neocerebellum à partir des mammifères.
- Subdivisions anatomiques sagittales:
- vermis;
- partie parasagittale des hémisphères cérébelleux ou paravermis;
- partie latérale des hémisphères cérébelleux.
- Subdivisions anatomiques longitudinales:
- Afférences: lobules de I à X
- fibres grimpantes issues des noyaux de l'olive bulbaire,
- fibres moussues issues de la moelle, du tronc cérébral et du cerveau.
- Efférences:
- faisceaux issus des noyaux cérébelleux profonds;
- faisceau issu du cortex cérébelleux vers les noyaux vestibulaires latéraux.
- Fonctions:
- motricité: coordination des mouvements, apprentissage supervisé et automatisation des mouvements (mémoire procédurale motrice), régulation du gain des réflexes;
- cognition et affectivité: contrôle des fonctions exécutives, végétatives et émotionnelles.
Anatomie
Vue extérieure
Le cervelet est composé d'une structure sagittale étroite, le vermis cérébelleux, flanqué de deux de chaque côté par deux hémisphère plus volumineux que lui. Ces hémisphères sont à leur tour subdivisés en une région parasagittale ou paravermis et une région latérale.
Trois cordons de substance blanche, appelés pédoncules cérébelleux, rattachent le cervelet à la face dorsale du tronc cérébral de chaque côtés :
- les pédoncules cérébelleux inférieurs issus des corps restiformes bulbaires ;
- les pédoncules cérébelleux moyens issus des fibres transverses du pont ;
- les pédoncules cérébelleux supérieurs qui convergent vers le tegmentum mésencéphalique.
Structure interne
Le cervelet se compose d'une substance grise périphérique, plissée (cortex à trois couches) qui circonscrit une substance blanche interne laquelle renferme
Des sillons concentriques dépriment la surface du cervelet et lui confèrent un aspect multi-folié sagittalement. Ces sillons définissent, en outre, par ordre décroissant de profondeur des lobes, lobules, lames et lamelles. Aussi le cervelet se divise-t-il en trois lobes principaux: antérieur, postérieur et nodulo-flocculaire, séparés par les sillons primaires et postéro-latéral, respectivement. À leur tour, ces lobes se subdivisent en dix lobules au total par des sillons secondaires.
Le cortex cérébelleux comprend un circuit fondamental, commun aux vertébrés tétrapodes, d'aspect quasi-cristallin et qui présente des propriétés de plasticité synaptique. Il s'agit d'un circuit à double entrée excitatrice et centré sur les cellules de Purkinje. La première entrée, disynaptique, est représentée par les fibres moussues qui se connectent aux grains lesquels, à leur tour, se connectent par les fibres parallèles à plusieurs cellules de Purkinje alignées parasagittalement. La seconde entrée est constituée par les fibres grimpantes qui se terminent sur les cellules de Purkinje. Sur ce circuit de base se greffent des micro-circuits inhibiteurs centrés sur six types actuellement connus d'inter neurones. Par exemple, les cellules stellaires contrôleraient les entrées des cellules de Purkinje, tandis que les cellules à paniers en contrôleraient les sorties. Les cellules de Purkinje se projettent sur les noyaux cérébelleux profonds ipsi-latéraux (hémisphère cérébelleux latéraux → noyau dentelé; paravermis → noyaux interposés; vermis→ noyau fastigial). Les axones des neurones de projection des noyaux cérébelleux profonds (et le noyau vestibulaire latéral) constituent les seules efférences du cervelet
Les trois couches du cortex cérébelleux
Le cortex cérébelleux comporte uniformément trois couches superposées:
- la couche moléculaire (immédiatement sous la pie-mère);
- la couche ganglionnaire; (couche des cellules de Purkinje)
- la couche granulaire, la plus interne.
- Couche granulaire
La couche granulaire est composée par les grains, neurones glutaminergiques et glycinergiques, qui reçoivent les fibres moussues au niveau de jonctions glomérulaires. Les grains émettent un axone ascendant épineux vers la couche moléculaire où il se divise en deux branches opposées, parallèles au grand axe du folium. Ces branches, appelées fibres parallèles, font synapses avec les dendrites des cellules de Purkinje. La couche granulaire comprend aussi des interneurones GABAergiques: les grandes et petites cellules de Golgi, et les cellules de Lugaro. Les cellules unipolaires à brosse (UBC) sont présentes au sein du cervelet vestibulaire (lobe flocculo-nodulaire, uvula, paraflocculus ventral) et au sein des noyaux cochléaires.
- Couche ganglionnaire
La couche ganglionnaire est formée par une juxtaposition monocellulaire des perikaryons piriformes des cellules de Purkinje GABAergiques dont les arbres dendritiques apicaux, aplatis, et épineux à partir de la troisième bifurcation, se distribuent en rangées para sagittales au sein de la couche moléculaire sus-jacente. Entre les cellules de Purkinje s'intercalent les cellules en candélabre.
- Couche moléculaire
La couche moléculaire renferme, outre les dendrites des cellules de Purkinje et des cellules de Golgi et les fibres parallèles, des inter neurones inhibiteurs GABAergiques : les cellules à paniers (majoritairement situées dans la moitié inférieure de la couche) et les cellules stellaires(majoritairement situées dans la moitié supérieure de la couche).
Afférences et efférences du cervelet
D'une manière générale, les fibres moussues font synapse avec les dendrites des grains au niveau des glomérules (et des cellules de Golgi) et les fibres grimpantes font synapse avec les cellules de Purkinje, mais ces deux types de fibres envoient une collatérale aux noyaux cérébelleux profonds.
- Afférences
- Afférence moussues:
-
- afférences spinales : Faisceaux spino-cérébelleux dorsal, spino-cérébelleux ventral, spino-cérébelleux rostral, et cunéocérébelleux ;
- afférences réticulaires : noyaux réticulaires latéral, paramédian, tegmental pontique ;
- afférences pontiques : noyaux pontiques, raphé, trigeminal, péri-hypoglossiens, du tractus solitaire, du vague, vestibulaires et parabracchial ;
- afférences mésencéphaliques : colliculi inférieurs et supérieurs ;
- afférences diencéphaliques : hypothalamus, pallidum.
Les noyaux pontiques situés dans la région ventrale de la protubérance (basis pontis) forment les principaux relais des afférences corticales vers le cervelet. Ces afférences comprennent, chez le singe, des faisceaux issus des cortex sensori-moteur et prémoteur, ainsi que des faisceaux "associatifs": prefronto-pontique (aires de Brodmann 8A, 9, 10, 45B, 46, 32), temporo-pontique (la berge supérieure du sillon temporal supérieur), pariéto-pontique (lobules pariétaux supérieur et inférieur), para-hippocampo-pontique, occipito-pontique (région parastriée),et cingulo-pontique (aires 23, 24, 25, 35 et cortex rétrosplénial).
- Afférence "grimpantes" :
- olive bulbaire principale et olives accessoires médiale et caudales.
L'olive bulbaire reçoit des afférences multiples via le faisceau cortico-bulbaire (cortex moteur), via le faisceau tegmental central (notamment la partie parvocellulaire du noyau rouge), via les noyaux cérébelleux profonds et via le faisceau spino-olivaire. Le noyau de l'olive bulbaire participe ainsi à une boucle cérébello-rubro-olivo-cérébelleuse dont le fonctionnement est modulé principalement par les projections corticales. Les fibres grimpantes se terminent sur les troncs primaires et secondaires des cellules de Purkinje, du côté opposé. À noter qu'une cellule de Purkinje est innervée par une seule fibre grimpante. Les fibres grimpantes induisent l'émission de potentiels d'action complexes et synchrones chez les cellules de Purkinje, et peuvent moduler le poids synaptique entre cellules de Pukinje et fibres parallèles. Cette modulation hétérosynaptique diminue globalement l'activité des cellules de Purkinje à court terme (pause ou sensibilisation) ou à long terme (dépression à long terme (DLT)). Cette DLT secondaire à une désensibilisation des récepteurs AMPA ionotropes serait à la base du processus d'apprentissage supervisé que sous-tendrait le cervelet. Ainsi, a-t-il été proposé que les fibres grimpantes délivreraient un signal d'erreur reflétant, par exemple, l'inadéquation entre le mouvement réalisé et le mouvement projeté, mais aussi que ces fibres participeraient à la détection de nouveauté, à la régulation du degré d'activation globale des cellules de Purkinje, à la synchronisation des cellules de Purkinje... Le ou les rôles exacts restent encore à déterminer.
Les neurones olivaires présentent une oscillation endogène de leur potentiel membranaire à une fréquence d'environ 10 Hz, et la survenue des potentiels d'action coïncide avec le maximum d'amplitude de ces oscillations. L'activité de plusieurs neurones olivaires peut être synchronisée par des jonctions communicantes au niveau de leurs dendrites. À l'opposé, les projections GABAergiques des noyaux cérébelleux concourent à désynchroniser le fonctionnement de ces neurones. L'activité de ces "agrégats" de neurones olivaires permettrait de séquencer dans le temps l'activité des cellules de Purkinje en définissant des fenêtres d'activation.
- Efférences
- des noyaux fastigiaux:
- faisceau unciné cheminant dans le pédoncule cérébelleux supérieur controlatéral et se terminant au niveau: de la formation réticulée, du raphé, des noyaux pontiques, des noyaux vestibulaires, de l'hypothalamus et du thalamus;
- faisceau fastigio-bulbaire direct cheminant dans le pédoncule cérébelleux inférieur ipsi-latéralement et se terminant sur les noyaux vestibulaires, olivaires accessoires, et la formation réticulée;
- faisceau fastigio-spinal : se terminant sur des motoneurones cervicaux contro-latéraux.
- des noyaux interposés:
- faisceau principal cheminant dans le pédoncule cérébelleux supérieur, décussant dans le tegmentum mésencéphalique ventral et bifurquant en une branche ascendante vers la partie magnocellulaire du noyau rouge, l'hypothalamus et le thalamus, et une branche descendante vers le noyau réticulaire tegmental pontique, les olives bulbaires accessoires et des interneurones medullaires.
- des noyaux dentelés:
- faisceau principal cheminant dans le pédoncule cérébelleux supérieur, décussant dans le tegmentum mésencéphalique ventral et bifurquant en une branche ascendante vers la partie parvocellulaire du noyau rouge, l'hypothalamus, la zona incerta et le thalamus, et une branche descendante vers le noyau réticulaire tegmental pontique, et l'olive bulbaire principale ;
- faisceau dentato-spinal : faisceau polysynaptique dont une partie fait relais dans la formation réticulée.
- faisceau principal cheminant dans le pédoncule cérébelleux supérieur, décussant dans le tegmentum mésencéphalique ventral et bifurquant en une branche ascendante vers la partie parvocellulaire du noyau rouge, l'hypothalamus, la zona incerta et le thalamus, et une branche descendante vers le noyau réticulaire tegmental pontique, et l'olive bulbaire principale ;
Les noyaux cérébelleux profonds se projettent aussi sous forme de fibres moussues glutaminergique ou, moins densément, GABAergiques sur les grains.
Les noyaux thalamiques recevant des projections cérébelleuses comprennent chez le singe: les noyaux ventro-postéro-latéral oral, ventro-latéral caudal, X, dorso-médian et intralaminaire (paracentral).
Par l'intermédiaire du thalamus, le cervelet se connecte, chez le singe, aux cortex moteur (aire 4), prémoteur (aire 6 incluant l'aire motrice supplémentaire), pariétal (aire 7) et préfrontal (aires 8, 9 latérale et médiane et 46 dorsale). Les voies cérébello-préfrontales sont issues de la région ventrale du noyau dentelé.

















































