Chimiosmose - Définition
La liste des auteurs de cet article est disponible ici.
Introduction
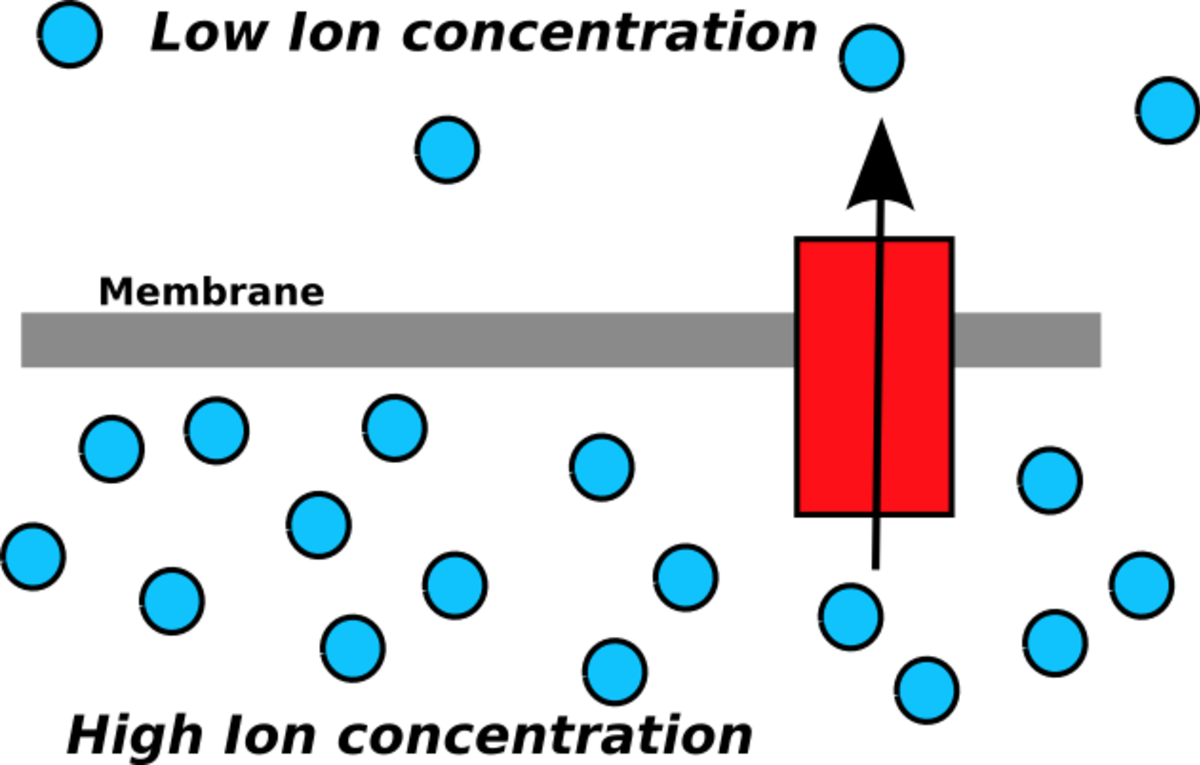
La chimiosmose est la diffusion des ions au travers d'une membrane sélectivement-perméable. Plus spécifiquement, cela concerne la production d'ATP par le déplacement des ions hydrogène au travers d'une membrane interne durant la respiration cellulaire.
Les ions hydrogène diffusent à partir de la zone ayant la concentration la plus élevée vers la zone ayant la concentration la plus basse. Peter Mitchell avança l'idée qu'un gradient électrochimique de protons à travers une membrane puisse être exploité pour produire de l'ATP. Il rapprocha ce processus de l'osmose, la diffusion de l'eau à travers une membrane, raison pour laquelle ce phénomène est nommé chimiosmose.
L'ATP synthase est une enzyme synthétisant l'ATP par chimiosmose. Elle permet le passage de protons à travers la membrane en utilisant l'énergie cinétique pour phosphoryler l'ADP en ATP. La production d'ATP par chimiosmose a lieu au niveau des chloroplastes et des mitochondries ainsi que chez quelques bactéries.
La théorie chimiosmotique
Peter D. Mitchell proposa l'hypothèse chimiosmotique en 1961. Cette théorie stipule essentiellement que la plupart de l'ATP synthétisée par respiration cellulaire provient du gradient électrochimique à travers les membranes internes des mitochondries par l'utilisation de l'énergie du NADH et de la FADH2 formés à partir de la réduction de molécules riches en énergie comme le glucose.
Des molécules comme le glucose sont métabolisées afin de produire de l'acétyl-CoA comme intermédiaire riche en énergie. L'oxydation de l'acétyl-CoA au sein de la matrice mitochondriale est couplée à la réduction de coenzymes comme la NAD et le FAD. Les coenzymes transfèrent les électrons à la chaîne de transport d'électrons (CTE) au niveau de la membrane mitochondriale interne qui à son tour les transfert à d'autres protéines de la CTE. Les apports d'énergie de ce transfert d'électrons sont utilisés pour pomper les protons de la matrice vers l'espace inter-membranaire, l'énergie étant ainsi stockée sous forme d'un gradient électrochimique transmembranaire. Les protons repassent ensuite la membrane interne au travers de l'ATP synthase. Le flux de protons retournant à la matrice mitochondriale via l'ATP synthase apporte assez d'énergie pour associer un phosphate inorganique à de l'ADP pour former de l'ATP. Les électrons et les protons de la dernière pompe de la CTE sont pris en charge par l'oxygène pour former de l'eau.
Ce fut une théorie radicale pour l'époque et elle ne fut pas admise : l'opinion commune était que l'énergie des transferts d'électrons était stockée sous forme d'intermédiaires stables à haut potentiel, un concept chimiquement plus conservateur.
Le problème avec cet ancien paradigme est qu'il n'y a jamais eu d'intermédiaires de haute énergie identifiés et que les preuves d'un pompage de protons par les complexes de la chaîne de transport d'électrons s'accumulèrent au point que cette théorie ne pouvait plus être ignorée. Finalement, aux vus de ces preuves, l'hypothèse chimiosmotique fut acceptée et, en 1978, Peter Mitchell reçu le Prix Nobel de chimie.
Le couplage chimiosmotique est important pour la production d'ATP au sein des chloroplastes et de quelques bactéries
Chez les mitochondries
La phosphorylation chimiosmotique est la troisième voie métabolique produisant de l'ATP à partir de phosphate inorganique et d'une molécule ADP. Ce processus fait partie de la phosphorylation oxydative.
La dégradation complète du glucose en présence de dioxygène est appelée la respiration cellulaire. Les dernières étapes de ce processus se déroulent dans les mitochondries. La réduction des molécules de NADH et de FADH2 se fait via le cycle de Krebs et la glycolyse. Ces molécules transfèrent les électrons à la chaîne de transport d'électrons qui utilise l'énergie créée par le gradient de protons de part et d'autres de la membrane interne. L'ATP synthase utilise ensuite l'énergie libérée par ce gradient afin de produire de l'ATP. Ce processus est appelée phosphorylation oxydative puisque le dioxygène est l'accepteur final d'électrons et que l'énergie libérée par la réduction du dioxygène en eau est utilisé pour phosphoryler l'ADP et produire de l'ATP.

















































