Évolution (biologie) - Définition
La liste des auteurs de cet article est disponible ici.
Mécanismes de l'évolution
L'évolution des populations
Parce que les individus d'une population possèdent des caractères héritables différents, et que seule une partie de ces individus accède à la reproduction, les caractères les plus adaptés à l'environnement sont préférentiellement conservés par la sélection naturelle. De plus, le hasard de la reproduction sexuée rend partiellement aléatoire les caractères qui seront transmis, par effet de dérive génétique. Ainsi, la proportion des différents caractères d'une population varie d'une génération à l'autre, conduisant à l'évolution des populations.
L'apparition de nouveaux caractères
Cela se produit par mutation et recombinaison génétique, ou remaniement chromosomique. Mais cela ne se déroule que dans un individu, pas dans l'espèce entière. Il faut, pour que ce nouveau caractère se répande, l'effet de la sélection naturelle et/ou de la dérive génétique.
Variabilité des individus au sein des populations
La plus part des individus d'une espèces sont uniques et diffèrent les uns des autres. Ces différences sont observables à toutes les échelles, du point de vue morphologique jusqu'à l'échelle moléculaire. Cette diversité des populations a deux origines principales: les individus sont dissemblables parce qu'ils ne possèdent pas la même information génétique et parce qu'ils ont subi des influences environnementales différentes.
La diversité génétique se manifeste par des variations locales de la séquence d'ADN, formant différents variants de la même séquence appelés allèles. Cette variabilité a plusieurs origines. Des allèles peuvent être formés spontanément par mutation de la séquence d'ADN. Par ailleurs, la reproduction sexuée contribue à la diversité génétique des populations de deux manières: d'une part, la recombinaison génétique permet de diversifier les combinaisons d'allèles réunies sur un même chromosome. D'autre part, une partie du génome de chaque parent est sélectionnée aléatoirement pour former un nouvel individu, dont le génome est par conséquent unique.
La diversité issue de l'environnement s'acquiert tout au long de l'histoire de l'individu, depuis la formation des gamètes jusqu'à sa mort. L'environnement étant unique à chaque endroit et à chaque moment, il exerce des effets unique sur chaque individu, et ce à toutes les échelles, de la morphologie jusqu'à la biologie moléculaire. Ainsi, deux individus possédant la même information génétique (c'est par exemple le cas pour les vrais jumeaux) sont tout de même différents. Ils peuvent notamment avoir une organisation et une expression différente de l'information génétique.
L'hérédité
Les êtres vivants sont capables de se reproduire, transmettant ainsi une partie de leurs caractères à leurs descendants. On distingue la reproduction asexuée, ne faisant intervenir qu'un individu, de la reproduction sexuée pendant laquelle deux individus mettent en commun une partie de leur matériel génétique, formant ainsi un individu génétiquement unique.
Les caractères génétiques, c'est-à-dire l'ensemble des séquences d'acide nucléiques d'un individu, ne sont pas tous transmis de la même manière. Lors de la reproduction asexuée, qui est une reproduction clonale, l'ensemble des séquences nucléiques sont copiées et l'information génétique contenue chez les deux descendants est alors identiques. En revanche, lors de la reproduction sexuée, il arrive fréquemment qu'une partie seulement du matériel génétique soit transmis. Chez les Métazoaires, les chromosomes sont fréquemment associés par paire, et seul un chromosome de chaque paire et de chaque parent est transmis à l'enfant. De plus, si les parents fournissent tous les deux la moitié du contenu nucléaire, le matériel cytoplasmique est souvent fournis par un seul des deux parents (la mère chez les mammifères). Ainsi, le matériel génétique contenu dans les organites semi-autonomes, tels que les chloroplastes et les mitochondries, n'est transmis que par une partie des individus de l'espèce (les femelles chez les mammifères).
La transmission des caractères acquis, une hypothèse non totalement rejetée
La théorie synthétique de l'évolution, paradigme dominant actuel, se fonde sur un déterminisme génétique intégral et écarte donc toute transmission héréditaire de caractères acquis au cours de la vie de l'individu. Néanmoins de plus en plus de travaux scientifiques remettent en cause ce modèle et rétablissent pour partie l'idée d'une transmission héréditaire de caractères acquis que défendait le lamarckisme.
Tout d'abord, certains caractères dits épigénétiques concernent la structure et l'organisation des génomes sont transmis par les parents en même temps que les molécules d'acide nucléique elles-mêmes. De plus, la mère fournit l'environnement cytoplasmique de la cellule-oeuf du descendant, et transmet ainsi un certain nombre de caractéristiques cellulaires à l'enfant. Des modifications épigénétiques conservées dans la lignée germinale sont désormais décrites chez plusieurs espèces. Chez les plantes il existe une corrélation entre le niveau d'expression d'un gène et sa méthylation. Pareillement, chez les mammifères nous témoignons de la méthylation d'une séquence transposable qui est insérée à proximité d'un gène particulier. Le degré de méthylation d'un transposon pouvant enfin moduler l'expression du gène dans lequel il s'est inséré. L'étude de l'épigénétique, longtemps délaissée, connait un grand essor depuis la fin du séquençage de nombreux génomes, dont celui de l'homme.
Ainsi, Une étude de 2009 du MIT affirme mettre en évidence une hérédité de certains caractères acquis chez des rongeurs. Par ailleurs, l’obésité serait non pas uniquement un effet direct touchant les individus atteints eux-mêmes mais également un effet transgénérationnel. Des données chez l'homme et chez l'animal semblent montrer que les effets d'une sous-alimentation subies par des individus pourraient en effet être transmises aux descendants.
La dérive génétique
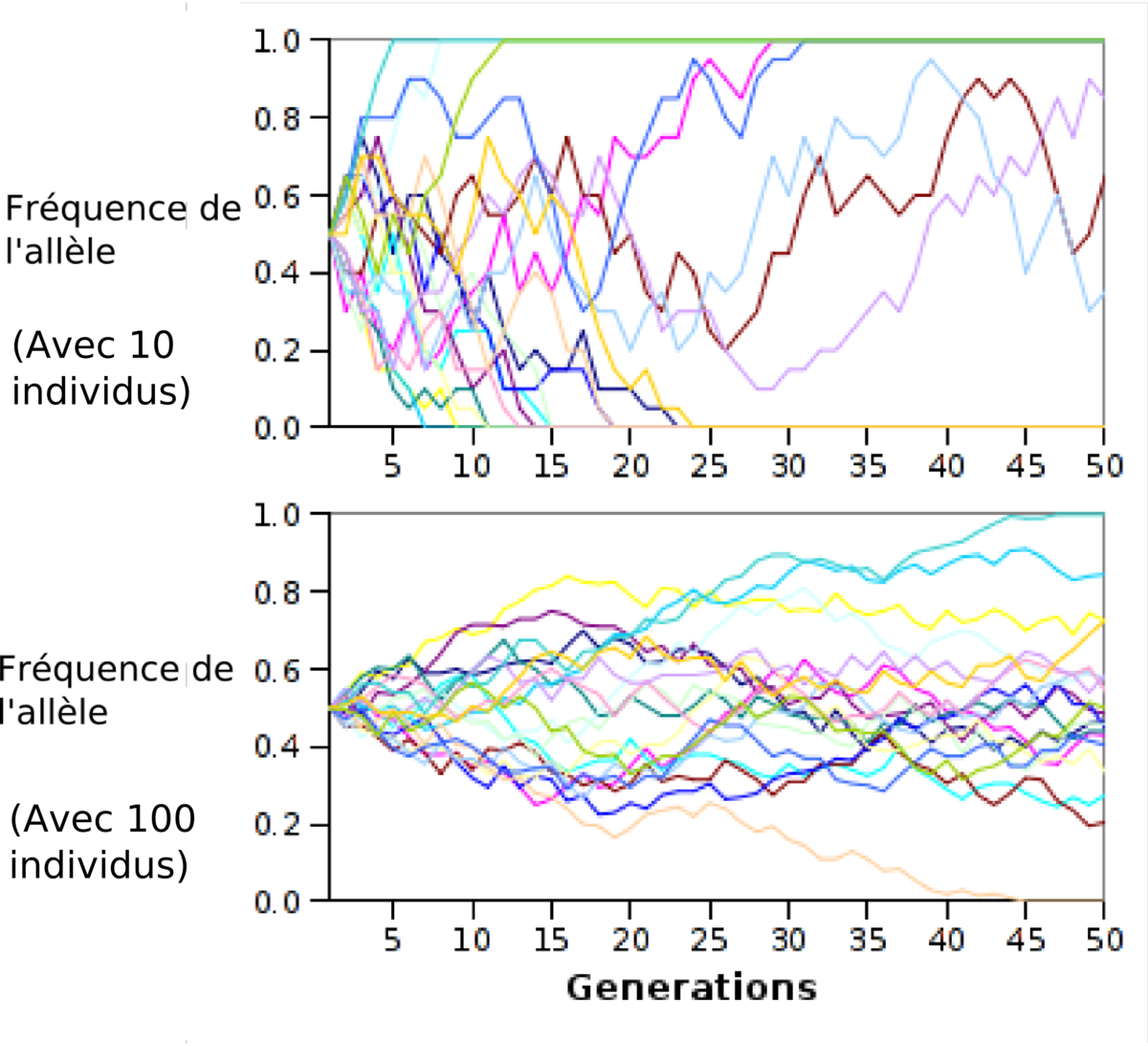
Lors de la reproduction sexuée, la transmission des caractères (notamment des allèles) comporte une grande part de hasard due à la recombinaison homologue, et au brassage génétique. Ainsi, on observe une variation aléatoire des fréquences alléliques d'une génération à l'autre, appelée dérive génétique. La dérive génétique génère donc une composante aléatoire dans l'évolution des populations. Ainsi, deux populations d'une même espèce n'échangeant pas de matériel génétique vont diverger jusqu'à former, si le temps d'isolement génétique est suffisant, deux espèces différentes. La dérive génétique est donc un des moteurs de la spéciation.
L'effet de la dérive génétique est particulièrement visible lorsqu'un faible nombre d'individus est à l'origine d'une population beaucoup plus nombreuse. C'est le cas lorsque se forme un goulot d'étranglement c'est-à-dire qu'une population est décimée et se reconstitue, ou lorsque quelques individus d'une population migrent pour aller coloniser un nouvel espace et former une nouvelle population (effet fondateur). Lorsqu'un tel évènement se produit, un allèle même faiblement représenté dans la population de départ peut se retrouver en forte proportion dans la population nouvellement formée sous le simple effet d'un hasard dans le tirage des individus à l'origine de la nouvelle population. Inversement, un allèle fortement représenté peut ne pas être tiré, et disparaît de la nouvelle population. Par ailleurs, la formation d'une nouvelle population à partir d'un faible nombre d'individu a pour effet d'augmenter la consanguinité dans la population et augmente le pourcentage d'homozygotie, ce qui fragilise la population.
La sélection naturelle
Dans la très grande majorité des espèces, le nombre de cellules-œuf produits est bien plus grand que le nombre d'individus arrivant à l'âge de la maturité sexuelle et parmi ceux-ci, une partie seulement accède à la reproduction. Ainsi, seule une partie des individus formés se reproduit à la génération suivante. Il existe donc une sélection des individus perpétuant l'espèce, seuls les individus n'étant pas éliminé par les conditions environnementales pouvant se reproduire. Cette sélection a été baptisée sélection naturelle.
Comme il existe une variabilité au sein des espèces, les individus possédant des caractères différents, et qu'une partie de ces caractères sont héréditaires, les caractères permettant à l'individu de survivre et de mieux se reproduire seront préférentiellement transmis à la descendance, par rapport aux autres caractères. Ainsi la proportion des caractères au sein des espèces évolue au cours du temps.
La sélection naturelle peut prendre des formes très variées. La sélection utilitaire est une élimination des individus les moins capables de survivre et les moins féconds, alors que la sélection sexuelle conserve préférentiellement les individus les plus aptes à rencontrer un partenaire sexuel. Bien que ces sélections soient complémentaires, on observe souvent des conflits, chaque forme de sélection pouvant favoriser l'évolution d'un caractère dans un sens différent.
Il est parfois observé une sélection d'individus qui favorisent la survie ou la reproduction d'individus qui leurs sont ou non apparentés, comme c'est le cas chez les insectes eusociaux ou lorsqu'un individu se sacrifie pour permettre la survie de son groupe ou de sa descendance. En sociobiologie, ces comportements altruistes s'expliquent notamment par les théories controversées de la sélection de parentèle, de la sélection de groupe et de l'altruisme réciproque. La sélection de parentèle prédit qu'il peut être plus avantageux pour un individu de favoriser beaucoup la reproduction d'un individu apparenté (donc avec lequel il partage des caractères) que de se reproduire un peu ou pas du tout, la sélection de groupe repose sur le même principe mais du point de vue du groupe et pourrait expliquer certains actes chez l'homme comme les guerres ou la xénophobie, l'altruisme réciproque se penche sur les cas d'altruisme entre individus non-apparentés et induit une contribution réciproque dont l'aide donnée en retour peut être différé dans le temps.
Enfin, la sélection artificielle n'est qu'une forme de sélection naturelle exercée par l'homme.
Conséquences évolutives
Adaptation des espèces
En conséquence de la sélection naturelle, les espèces conservent préférentiellement les caractères les plus adaptés à leur environnement, et sont donc de mieux en mieux adaptées à leur environnement. Les pressions de sélection en jeux dans cette adaptation sont nombreuses et concernent tous les aspects de l'environnement, des contraintes physiques jusqu'aux espèces biologiques interagissantes.
L'adaptation de plusieurs espèces différentes sous l'effet des mêmes pressions environnementales peut conduire à l'apparition répétée et indépendante du même caractère adaptatif chez ces espèces, par un phénomène de convergence évolutive. Par exemple, chez les mammifères les cétacés et les siréniens ont tout deux développé des nageoires, de manière indépendante. L'évolution de ces nageoires montre une adaptation convergente à la vie aquatique.
Cependant, l'effet de la sélection naturelle est réduit par celui de la dérive génétique. Ainsi, un caractère avantageux pourra ne pas être sélectionné à cause de l'inertie donnée par la dérive.
Apparition et disparition des espèces
L'évolution d'une population sous l'effet du hasard et des contraintes environnementales peut aboutir à la disparition de la population et éventuellement de l'espèce à laquelle elle appartient. Inversement, deux populations peuvent s'individualiser au sein d'une même espèce jusqu'à former deux espèces distinctes par un processus nommé spéciation.
Controverses sur les mécanismes de l'évolution
L'évolution et ses mécanismes sont encore largement étudiés aujourd'hui, et de nombreux points sur les mécanismes de l'évolution ne sont pas éclaircis. Certaines questions déjà soulevées par Charles Darwin n'ont d'ailleurs toujours pas de réponse certaine.
Une des grandes questions de la théorie de l'évolution est l'origine des rangs taxinomiques supérieurs à celui de l'espèce. En outre, la manière dont sont apparus les 33 embranchements animaux, issus de l'explosion cambrienne, pose encore problème. Ainsi, la théorie gradualiste estime que les changements interviennent de manière progressive au cours de l'évolution, alors que la théorie des équilibres ponctués défend qu'il existe des sauts évolutifs majeurs.
La transmission des caractères acquis, complètement délaissée depuis la découverte des lois de l'hérédité, est réactualisée par la découverte des phénomènes épigénétiques. Dès lors, l'importance de cette transmission de caractères non hérités des parents dans l'évolution des espèces doit se poser. Cependant, notre connaissance des mécanismes épigénétiques est encore trop faible pour pouvoir répondre à cette question. En outre, peu d'études sur le rôle de l'épigénétique dans l'évolution ont été réalisées à l'heure actuelle.
Il a été longtemps admis que l'évolution s'accompagnait d'un accroissement de la complexité des êtres vivants. Cependant, cette idée, largement influencée par l'anthropocentrisme, est fortement débattue aujourd'hui. La complexité n'ayant pas de définition précise à l'heure actuelle, il est difficile de vérifier une éventuelle augmentation de complexité. Par ailleurs, lorsque cette idée est admise, les origines de cette augmentation de complexité sont, elles aussi, source de controverse. En fait, tout cela à déjà été clairement expliqué par Lamarck.