Cetacea - Définition
La liste des auteurs de cet article est disponible ici.
Description
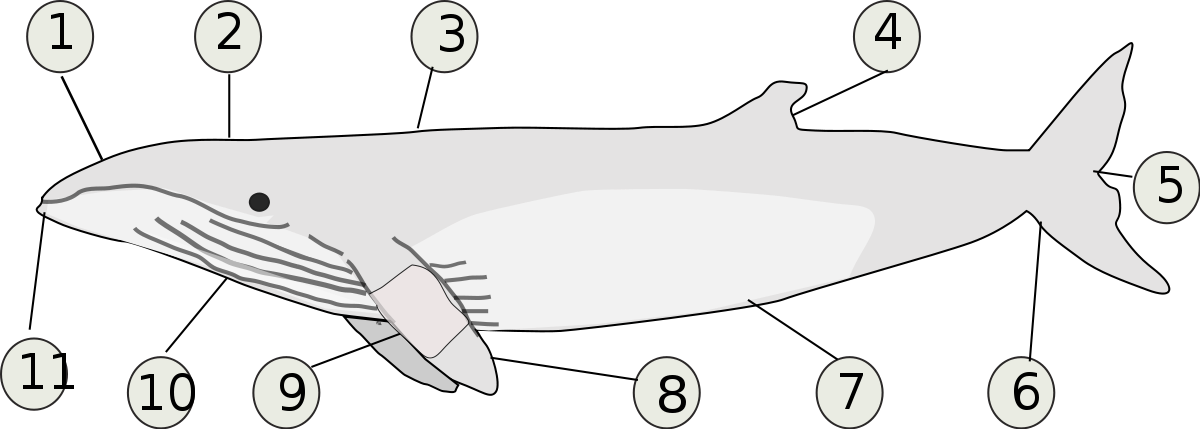
Baleine crème
1. Mâchoire supérieure
2. Évent
3. Peau noire, blanche ou grise
4. Nageoire dorsale en forme de faucille
5. Entaille médiane
6. Nageoire caudale
7. Dessous gris pale ou blanc
8. Les nageoires ont des points
9. Bande blanche sur chaque nageoire
10. 50 à 70 sillons ventraux
11. Museau à points
Morphologie
Les cétacés partagent tous un certain nombre de caractères morphologiques, un corps lisse et fuselé, ce qui facilite la pénétration dans l'eau, un système pileux quasi inexistant, l'absence de griffes, pas d'oreilles externes et le conduit auditif est fermé. Les membres antérieurs et la queue des cétacés ont évolué en nageoires. La nageoire caudale appelée palette, très puissante, est horizontale, contrairement à celles des poissons. Celle-ci assure la propulsion tandis que les nageoires latérales assurent l'équilibre et le maintien de la direction. L'aileron dorsal fait office de stabilisateur comme la quille d'un bateau. Comme les autres mammifères, les cétacés respirent à l'aide de poumons et doivent régulièrement faire surface pour respirer avec leur évent situé au sommet du crâne.
Le dimorphisme sexuel observé pour ces espèces est principalement lié à la taille, les mâles odontocètes sont en général plus grands que les femelles, mais ce n'est pas le cas par exemple chez les botos et chez la plupart des mysticètes dont la baleine bleue. Les mâles de certaines espèces d'odontocètes, comme les Ziphiidae ou surtout le narval, possèdent une dentition différente des femelles.
La livrée des cétacés est adaptée au mode de vie de l'animal. Elle peut être asymétrique gauche-droite comme pour le rorqual commun. Pour plusieurs espèces, elle permet de reconnaître les spécimens. La plupart des cétacés océaniques ont le dos foncé et le ventre blanc, de façon à ce que les proies et les prédateurs les confondent avec le fond de la mer lorsqu'ils les voient de dessus et ne voient qu'un reflet de la surface de l'eau lorsqu'ils les voient par dessous.
Des caractéristiques morphologiques permettent de reconnaitre les espèces, voire les individus, cependant l'observation des cétacés est difficile, du fait de leur vitesse et de leur méfiance vis-à-vis de l'homme. Les analyses génétiques permettent de résoudre cette difficulté : en effet, à chaque plongeon les cétacés laissent des squams, des résidus huileux ou des excréments qui flottent. Ceux-ci peuvent être analysés. Les espèces peuvent également être identifiées par leurs chants, toutes les espèces ne produisant pas des vocalises identiques.
Les spécimens de certaines espèces peuvent être facilement reconnus par les taches sur leur peau. Ainsi les spécimens de certaines espèces peuvent être reconnus uniquement par les taches sur leur nageoire caudale. Les baleines les plus grandes peuvent être aussi reconnues par la forme de l'implantation des balanes sur leur peau ou la forme de leurs nageoires. Les marques sur la peau, souvent présentes chez les mâles car résultant de combats, permettent aussi d'identifier les individus.
Selon Alexey V. Yablokov, la livrée des cétacés est liée à leur mode de vie, ainsi les cétacés aux livrées plus ou moins uniformes seraient planctophages, les livrées des espèces au ventral dégradé clair seraient sociales et les espèces aux livrées contrastées seraient des chasseurs.
| Désignation | Mysticètes | Odontocètes |
|---|---|---|
| Évent | Ouverture nasale double | Ouverture nasale unique |
| sillons ventraux | présent pour quelques espèces | absent |
| rostre | absent | plus ou moins long selon les espèces |
Les nageoires

Les cétacés possèdent deux nageoires pectorales ou palettes natatoires thoraciques, une nageoire dorsale ou aileron et une nageoire caudale ou queue. Les nageoires caudales permettent aux cétacés de s'orienter. Il semble que la nageoire dorsale permettent une plus grande efficacité dans les mouvements. Les nageoires dorsales de certains épaulards mâles peuvent dépasser deux mètres. Megaptera novaeangliae dispose quant à elle des plus grandes nageoires pectorales, celles-ci peuvent atteindre le tiers de leurs corps.
La progression est due aux pressions sur l'eau d'une nageoire caudale de forme homocerque. Celle-ci est orientée horizontalement, alors qu'elle est verticale chez les poissons osseux et cartilagineux. La pression de la queue sur l'eau est exercée verticalement, le sens de mouvement de cette nageoire favoriserait les déplacement de ces animaux du fond vers la surface et inversement. Le marsouin de Dall, un des cétacés les plus rapides, a été mesuré à 55 km/h, l'orque pourrait l'être encore plus. Hormis les siréniens, les autres mammifères marins ne possèdent pas de queue qui permette le mouvement.
Les sens
Le sens le plus développé chez les cétacés est l'audition, particulièrement chez les cétacés à dents qui sont dotés de la fonction d'écholocation, sorte de sonar naturel. Ils émettent des clics d'écholocation dont une partie est réfléchie par les obstacles. L'analyse de ces retours leur donne une sorte d'image acoustique de leur environnement. La présence de ce type de sens est beaucoup moins développée chez les baleines à fanons, mais des signes semblent montrer qu'elles n'en sont pas dépourvues.
Leur vue n'est pas mauvaise, même si elle n'est pas leur sens principal. Elle est beaucoup plus mauvaise pour les dauphins de rivières, certaines espèces sont même presque aveugles, pour les autres, elle est efficace sous l'eau ou hors de l'eau. Certains cétacés sortent la tête de l'eau pour observer les oiseaux et en déduire où se trouvent les bancs de poissons. Les cétacés des delphinariums sont capables de localiser des objets à plusieurs mètres au-dessus du niveau de l'eau et de s'en saisir. Les odontocètes semblent avoir perdu leur odorat, et il est vraisemblablement très réduit chez les mysticètes. Les globes olfactifs du cerveau ne semblent présent que chez les embryons. D'ailleurs, les chémorécepteurs comme l'organe de Jacobson sont atrophiés chez les Mysticètes et absents chez les Odontocètes.
Ce sens est compensé par un sens du goût, ainsi les grands dauphins sont capables de détecter par exemple des phéromones, des poissons... Ce même dauphin, s'il semble posséder moins de récepteurs chimiques permettant le goût, il est, en revanche, plus sensible au variation de salinité et surtout d'amertume.
Si toutes les espèces naissent avec des poils sur leurs rostres, seules certaines espèces de Mysticètes gardent des vibrisses à l'âge adulte. En revanche toutes ces espèces ressentent les contacts, la chaleur, leur position relative dans l'espace.
Une hypothèse couramment admise mais non prouvée suppose que les cétacés possèderaient un sens magnétique analogue à celui des oiseaux ou des tortues marines. Ceux-ci l'utiliseraient pour migrer sur de longues distances. La présence de magnétite à l'intérieur de leur cerveau corrobore cette hypothèse. Les relevés par satellite des baleines à bosse au large de l'archipel d'Hawaii montre que les trajectoires de ces dernières suivent le nord magnétique avec une grande précision. Certains expliquent par des erreurs dues à ce sens, les échouages, régulièrement observés depuis l'Antiquité au moins.
| Sens | Mysticètes | Odontocètes |
|---|---|---|
| écholocation | soupçonnée | présente |
| odorat | absent | soupçonné ou très réduit |
| sens magnétique | inconnu | soupçonné |
Les organes et leurs fonctions
Les ancêtres des cétacés, en quelques millions d'années se sont adaptés du milieu terrestre au milieu marin. À partir d'une morphologie terrestre proche des artiodactyles, ils ont perdu certaines caractéristiques pour en gagner d'autres. Comme chez les ruminants et autres ongulés, les cétacés possèdent trois bronches, trois poches stomacales, pas de clavicule, etc. Mais les adaptations au milieu aquatique comme celle du squelette et surtout les adaptations liées aux sens et celles liées aux capacités hyperbares pour certaines espèces sont très originales. On peut tout de même citer l'absence de glande sébacée, la présence d'une couche de graisse importante, les poumons et le foie non lobés, un diaphragme oblique... L'étude de l'anatomie des grandes espèces est particulièrement malaisée du fait de la taille imposante des animaux et donc des organes, le cœur d'une baleine bleue adulte pèse plus de 500 kilogrammes.
Les nageoires pectorales sont le résultat de l'évolution des membres supérieurs des cétacés, à ce titre, elle contiennent des os internes qui rappellent les os des pattes des mammifères. Les autres nageoires, dorsale ou caudale, sont en revanche dépourvues d'os et de muscles mais sont constituées de tissus fibreux rigides extrêmement solides. Les tissus de la nageoire caudale sont enveloppés de ligaments inextensibles et très solides attachés aux vertèbres.
Le génome des cétacés est diploïde, ils ont 42 ou 44 chromosomes.
Le système respiratoire et circulatoire

Le système respiratoire et circulatoire des cétacés est remarquablement efficace. Il doit répondre à deux objectifs pour la majorité des espèces, à savoir assurer des plongées les plus longues possible et de supporter des pressions élevées. Les performances mesurées au sein de ce taxon sont cependant très variées, les dauphins dulçaquicoles n'ont en effet pas besoin de descendre très profondément et donc pas besoin de plonger longtemps. La plus longue plongée enregistrée pour un Sousouc n'est que de 180 secondes ainsi que de 460 secondes pour une Orque mais de 3780 secondes pour une Baleine boréale, soit 21 fois plus longtemps que pour le sousouc.
La trachée et les bronches sont séparées, ce qui implique que les cétacés ne peuvent respirer que par les évents. La respiration est contrôlée et volontaire et non pas réflexe, si l'animal est anesthésié sans ventilation pulmonaire active ou ses évents bouchés, il meurt. Ils peuvent respirer relativement vite, un Grand dauphin respire trois fois par minute. L'évent est partiellement fermé pendant les expirations et grand ouvert pendant les inspirations. Les expirations expulsent de l'air chaud, mêlé de vapeurs, ce qui en région froide, produit des panaches visibles de loin à cause de la condensation de l'eau. Les cétacés utilisent 80 à 90% de leur volume pulmonaire, contre 10 à 15% environ pour l'homme, ce qui leur permet d'extraire 12% de l'oxygène qu'ils inhalent contre 4%, classiquement chez les mammifères terrestres.
Le volume sanguin des cétacés est élevé puisqu'il représente 15 % du poids de leur corps, contre 7 % chez les humains. En outre, il dispose de deux fois plus d'hématies et de myoglobines que ces derniers. Ils sont également capables de contrôler l'alimentation en oxygène des organes non vitaux comme les intestins. La fréquence de battements de cœur de certains d'entre eux baisse à 80 par minute en plongée.
Le Rete mirabile est bien développé comme pour tous les mammifères marins. Il l'est surtout dans la cage thoracique où il forme un plexus sous la plèvre, entre les côtes et de chaque côté de la colonne vertébrale. Il est également développé dans l'aire à la base du crâne. Les dispositions et mécanismes des veines et artères autour du cœur, qui est relativement aplati, ont pour but de ralentir le retour du sang veineux et donc de réduire la fréquence de battement.
En général, les mammifères marins ont proportionnellement des plus petits poumons que les autres mammifères, ceux des cétacés sont particulièrement élastiques et placés dorsalement dans la cage thoracique. Ils reposent sur un diaphragme plus long et placé plus horizontalement que chez les autres mammifères. Ils plongent donc avec relativement peu d'air en comparaison de leur volume total. Il plongent d'autant plus facilement que leur masse volumique est plus élevée sans un grand volume d'air. Les humains qui plongent en utilisant des bouteilles, malgré leur lourd équipement, doivent ajouter des plombs à leurs accessoires pour arriver à descendre. Lors de la descente, l'air vicié repasse du sang aux poumons, évitant aux gaz de stationner dans le système sanguin. Ils expirent donc dans l'eau lors de la plongée pour évacuer les surplus de gaz. Il semblerait que les Grands Cachalots secrètent également une « huile » fixant l'azote du sang. Ainsi, ils ne craignent donc ni l'ivresse des profondeurs et ne sont que rarement victimes d'accidents de décompression.
Comme chez les plongeurs professionnels humains, on peut observer une ostéonécrose c'est-à-dire une nécrose des tissus cartilagineux au niveau des articulations. C'est une conséquence de l'accumulation de l'azote qui provoque de minuscules accidents de décompression. Certains experts estiment que les grands cachalots qui peuvent plonger à plus de 2 500 mètres pendant plusieurs heures observent des paliers de décompression.
Les muscles et le squelette
Le squelette

Les modifications au niveau du squelette sont nettement visibles et sont analogues sur plusieurs points à celles des siréniens, mais il s'agit d'une convergence évolutive. Le rôle du squelette, n'est pas de porter le corps, mais de servir de support aux ligaments. Le poids moyen de leur squelette a été évalué à 17% soit un peu plus que ceux des hommes, évalué à 15%, mais moins que ceux des éléphants, évalué à 20%.
Les membres pelviens sont considérablement réduits : il ne reste plus que quelques os pelviens résiduels auxquels est relié le muscle ischio-caverneux qui maintient le pénis en place à l'intérieur de la fente génitale chez les mâles. Ce muscle est analogue à celui retrouvé chez les autres mammifères dont, entre autres, les ongulés et l'Homme. En revanche, les membres thoraciques sont conservés, quoique atrophiés, et l'on retrouve la Scapula (« omoplate » de l'Homme), l'humérus, le radius, l'Ulna (anciennement « cubitus »), les os du Carpe, les métacarpes et les phalanges qui se sont multipliées, on parle d'hyperphalangie. Les membres supérieurs ne sont pas mobiles les uns vis-à-vis les autres à part pour l'épaule. La clavicule est absente comme elle l'est chez les autres ongulés. Le tout forme une nageoire pectorale, équivalente à celle des Poissons et dont le rôle n'est pas la propulsion mais le maintien de l'équilibre dans l'espace et la direction des mouvements. Les autres nageoires ne sont pas ossifiées.
Parmi les autres caractéristiques observées, on observe un cou court et peu flexible, les vertèbres sacrées ne sont pas fusionnées.
C'est au niveau du crâne que l'on différencie principalement les Odontocètes des Mysticètes, les mysticètes ont un crane symétrique, pas les odontocètes. Les os de la face (maxillaire, incisif, mandibule, etc...) se sont allongés et se télescopent avec les os du crâne sur la face dorsale du crâne pour former ce que l'on appelle l'évent, issu de la migration sur la face dorsale des narines osseuses au cours de l'Oligocène.
Les côtes sont très fines voir absentes, sauf chez les Baleine pygmée. Cette adaptation confère un avantage important pour les plongées profondes. Le nombre de phalanges dépend des espèces mais est toutefois important, permettant de rigidifier les nageoires pectorales de grande taille.
Il n'y a pas d'os dans les nageoires dorsale et caudale en revanche, les nageoires pectorales en contiennent et ils sont les homologues de ceux, par exemple, de la main humaine. De très rares spécimens disposent d'embryons de nageoires à moitié arrière du corps, parallèlement au nageoires pectorales, ce sont, selon la théorie de l'évolution, des vestiges des membres arrières, peu à peu atrophiés au fil des générations.
| Mysticètes | Odontocètes |
|---|---|
| Absence de dents (sauf à l’état embryonnaire), présence de fanons | Présence de dents, quelques fois présentes simplement dans la gencive |
| Crâne symétrique | Crâne asymétrique |
| Absence de côtes sternales | Présence de côtes sternales |
| Sternum composé d'un os unique qui s’articule uniquement sur la première paire de côte | Sternum composé de trois os s’articulant avec trois paires de côtes ou plus |
L'attache des muscles dorsaux au squelette varie énormément entre les différentes familles, c'est un des critères possibles d'identification de celles-ci.
Les muscles
La chair est particulièrement foncée, car les muscles sont riches en myoglobine. La musculature est particulièrement développée puisqu'elle représente 40% de la masse corporelle de la Baleine bleue et 54% du Rorqual boréal. L'organisation de la musculature est foncièrement différente de celle des autres mammifères, les muscles peauciers peuvent être réduit mais leur masse peut être importante.
Le système tégumentaire
- Les fanons des mysticètes sont fixés sur les mâchoires supérieures, les plus long fanons des plus grandes espèces sont supérieurs à 1,5 mètres de large. Ces fanons leurs servent à filtrer ce que contient leur bouche en expulsant l'eau.
- Les cétacés sont homodontes c'est-à-dire que leurs dents, sauf exception comme le narval qui en principe ne possède qu'une défense, sont identiques entre elles. La denture est très différentes parmi les espèces de ce groupe, mais les embryons de toutes ces espèces disposent de dents. Les mysticètes perdent ces dents avant leur naissance, tandis que certains espèces de Platanistidae en ont plus de 120. Les dents de lait des odontocètes ne tombent pas, la seconde série reste atrophiée. Les dents des odontocètes servent à agripper les proies et leur forme dépend de leur régime alimentaire. Ainsi, leur nombre et leur forme permet aux spécialistes d'en déduire l'espèce. Les cétacés avec des dents sont plutôt piscivores, ceux sans dent sont plutôt consommateurs de céphalopode.
- poils sensitifs sont fixés sur les commissures de la bouche pour les mysticètes.
| Mysticètes | Odontocètes |
|---|---|
| présence de fanons | présence de dents, au moins deux |
| présence de poils | absence de poils |
La peau
La peau est composée de quatre couches dont les épaisseurs sont différentes de celle des autres mammifères. La couche épidermique des cétacés atteint 2 à 4 millimètres en moyenne, et 12 millimètres chez le bélouga. L'épiderme est composé de tissu conjonctif dense et est en général dépourvu de follicules pileux et de glandes sébacées. Certaines zones, pour certaines espèces, sont encore équipées de poils sensibles. La couche épidermique est fine, lisse et élastique et exsude de l'huile qui augmente leur hydrodynamisme. Cette adaptation est également présente chez de nombreux poissons. Ainsi les Mysticètes peuvent atteindre des vitesses de 26 km/h et les odontocètes des vitesses supérieures à 30 km/h. Le paradoxe de James Gray, qui stipule que les muscles des cétacés sont insuffisamment puissants pour permettre à ceux-ci de se déplacer aux vitesses observées est aujourd'hui caduc, les muscles le sont. Plusieurs hypothèses pour expliquer l'inadéquation musculaire supposée à leur vitesse réelle ont été formées. L'une d'elle proposait qu'il se formait à la surface de la peau une multitude de rides minuscules qui dirigent le flux de l'eau en évitant la création de micro-turbulences qui freineraient ces animaux, permettant de conserver un écoulement régulier de l'eau favorisant un glissement dans l'eau avec peu d'efforts.
Le derme est composé de tissus conjonctifs d'une épaisseur formée par quelques cellules. L'hypoderme est lui plus épais, il est poreux et inextensible. La concentration en graisse y est importante, ainsi l'hypoderme de la Baleine Bleue est constitué de 62% de graisse soit 15 à 20 tonnes au total. La proportion en graisse varie entre les couches extérieures et les couches intérieures. La quantité de graisse varie elle en fonction des saisons, des migrations, de la quantité de nourriture disponible. Comme chez les autres mammifères, cette couche sert de réserve énergétique. C'est dans cette couche que l'on retrouve les grandes accumulations de polluants auxquels sont soumis les cétacés. Cette couche contient des tissus conjonctifs intermédiaires et est très vascularisée. Ce système permet aux cétacés de contrôler leur température interne. La quatrième couche, le Stratum germinativum est constitué d'un Épithélium pavimenteux et les mélanocytes y sont fixés.
La peau des cétacés est aussi sensible au toucher.
Le système uro-digestif
Osmorégulation
L'eau de mer est un milieu Hypertonique, c'est-à-dire que les cellules des organismes marins doivent sans cesse lutter contre la perte naturelle de leur eau. Cette lutte nécessite une adaptation au moins aussi poussée que celle des mammifères vivant dans les zones très arides. Il existe assez peu d'études sur ce sujet pour ces animaux.
La concentration en sel dans leur urine est faible de tel sorte qu'elle est plus peu être plus faible que la salinité ambiante oblige à se poser la question de ce qu'ils boivent. Les cétacés en boivent cependant très peu, selon une étude de 1970, entre 4,5 et 13 ml par kg et par jour. Ces espèces ne règlent pas leur concentration interne en sels par l'absorption de l'eau de mer.
Seul parmi les mammifères, les reins des cétacés, des pinnipèdes, des loutres ou des ours ont une structure réniculée. Ces reins sont constitués d'une multitude de petits lobes qui contiennent des tissus corticaux et une pyramide rénale insérée dans un seul calice. La structure des reins ne permet pas d'expliquer leur performance, la raison de leur performance est vraisemblablement due au mécanisme de régulation hormonale de la concentration d'urine.
Le système digestif
Les Cétacés sont carnivores pourtant leur système digestif ressemble à ceux des Ruminantia, il possède trois poches. Ils n'ont pas de vésicule biliaire. Le foie est non lobé.
La première poche stomacale est un assez grand et musculeux estomac mécanique qui broie les aliments. Cela est d'autant plus nécessaires que leurs dents, quand ils en ont, ne sont pas destinées à broyer la nourriture. Le seconde poche est la plus grande, les sucs digestifs commence à dissoudre la nourriture. L'estomac pylorique est le plus petit des trois estomacs, commence là l'assimilation des substances nutritives. Cet estomac est suivi d'une ampoule duodénale, puis du duodénum. L'élimination des sels et donc les reins jouent un rôle parallèle important.
Les organes reproducteurs
Les organes génitaux sont internes, situés derrière la fente génitale, chez les mâles comme les femelles si bien qu'il est parfois difficile de distinguer le sexe des spécimens. Les testicules des mâles sont également dans cette poche. L'utérus des femelles est constitué de deux parties comme chez les autres ongulés et les carnivores, les mamelles, plates, élongées et extractibles, se trouvent près de la fente génitale. Le placenta est épithéliochoriale.
Chez les femelles des cétacés, contrairement aux autres mammifères, les corps blancs ne disparaissent pas et témoignent de l'histoire reproductive de chaque animal. Les cétologues peuvent ainsi estimer le nombre de gestations d'un cétacé.
La thermo-régulation
La graisse
La couche de graisse sous-cutanée, généralement très épaisse, permet d'éviter la déperdition de chaleur, épouse la forme du corps et équilibre le poids de ce dernier, augmente la flottabilité. La Pannicule de certaines baleines peut atteindre 30 cm.
Le système nerveux et sens
L'oreille et l'ouïe
Les cétacés n'ont plus d'oreilles externes mais se servent, en plus des osselets habituels aux mammifères, de petits os dans la mandibule qui transmettent les sons grâce au coussin adipeux qui entoure l'os tympanique. L'oreille interne est isolée par une ampoule contenant de l'air. Les variations de densité de toutes ces structures indiquent à l'animal d'où vient le son. Le nombre de cellules qui innervent l'oreille - deux ou trois fois plus élevé que chez l'homme - et la taille importante du nerf auditif indiquent que les cétacés parviennent très bien à discriminer les tons et les ondes sonores à haute fréquence et surtout le lieu de leur provenance.
La production des sons
Les cétacés peuvent produire un grand nombre de son destinés à communiquer comme la production de bulles, de sifflements, de bruit de battements de nageoire, de vocalises et de trombes avec leur évents, mais pas seulement pour communiquer. Le sonar nécessite l'émission de cliquetis pour fonctionner. Le Grand Cachalot et l'Épaulard ainsi que quelques espèces de dauphins sont capables d'étourdir et de paralyser les poissons et les calmars avec de puissantes ondes sonores.
Les yeux et la vue
Les cétacés n'ont pas de paupières, leurs yeux secrètent une huile qui les protège. Ils n'ont pas de glandes lacrymales. Leurs yeux sont souvent petits par rapport à leur taille en comparaison de ceux des primates par exemple. La sclère est très épaisse, particulièrement à l'arrière de l'œil dans la zone du nerf optique ce qui réduit encore le volume du globe oculaire et donc l'acuité visuelle. Le cristallin est presque sphérique.
L'arrière de la rétine est constitué d'un tapetum lucidum, de type cellulosum choroïdien, une zone richement vascularisée qui permet d'accroitre la quantité de lumière qui se dirige vers la rétine, autrement dit cela accroit la vision dans l'obscurité. La rétine contient des cônes et des bâtonnets, ce qui leur permet de détecter les couleurs, mais les cônes ne représentent que 1% des photo-récepteurs et ceux sensibles aux faibles longueurs d'onde ont disparu. Bien que cette question soit sujette à controverse, la vision des couleurs ne leur est certainement possible que dans certaines circonstances.
Les yeux sont disposés latéralement ce qui rend la vision stéréoscopique impossible pour certaines espèces. Les Delphinidae peuvent bouger, tel les caméléons leurs yeux indépendamment. Les grands dauphins peuvent même focaliser leurs yeux et les zones de chevauchement leur permettraient de percevoir les objets en relief.
Mais, même si ce n'est pas leur organe sensoriel principal, ils n'en sont pas pour autant atrophiés. Le grand Dauphin sait même adapter sa vision hors de l'eau de sorte qu'en observant le vol des oiseaux de mer, il peut repérer les bancs de poissons.
Le cerveau
Le cerveau des cétacés est très performant. Avec les éléphants et les primates, les cétacés font partie des rares animaux dont la conscience d'eux-mêmes a été scientifiquement mise en évidence. Les expériences sur le Grand dauphin montrent qu'ils sont également capables de se mettre à la place d'un humain, d'apprendre, d'inventer mais ils ont également conscience de leurs propres états de la connaissance (métacognition). D'autre part, on sait que les cétacés sont capables de communiquer, que tous les groupes d'une même espèce n'utilisent pas forcément les mêmes signaux, y compris vocaux, pour communiquer, de maitriser une grammaire et même, pour certaines espèces au moins, de pouvoir nommer par vocalise un partenaire. Le cerveau des cétacés a évolué alors qu'ils étaient déjà mammifères marins, d'une façon indépendante de celui des primates depuis 95 Ma et il est donc remarquable qu'il permette des performances comparables à celles des primates sur ces points. C'est un remarquable exemple de convergence évolutive.
En fait le néocortex des cétacés surpasse par son degré de gyrification celui de tous les autres mammifères, y compris des humains. L'épaisseur des différentes couches du néocortex diffère cependant de celle des primates, la I et la IV sont plus minces, la II plus dense. En fait la connectivité intra-hémisphérique semble favorisée chez les cachalots par exemple. Le cerveau des cétacés se caractérise aussi par la forte densité des cellules gliales. Ceci rend les cerveaux des cétacés très différents de ceux des espèces terrestres même si, curieusement les corticaux préfrontaux dédiés aux fonctions cognitives élevées comme l'attention, le jugement, l'intuition, et la conscience sociale, sont à la même position que ceux des primates.

















































